
Tăiați aici exemplele dumneavoastră.
- 9.1 Molecule de semnalizare și receptori celulari
- 9.2 Propagarea semnalului
- 9.3 Răspunsul la semnal
Imaginați-vă cum ar fi viața dacă dumneavoastră și persoanele din jurul dumneavoastră nu ați putea comunica. Nu ați putea să vă exprimați dorințele față de ceilalți și nici nu ați putea pune întrebări pentru a afla mai multe despre mediul în care vă aflați. Organizarea socială este dependentă de comunicarea dintre indivizii care alcătuiesc acea societate.
Ca și în cazul oamenilor, este vital pentru celulele individuale să poată interacționa cu mediul lor și între ele. Acest lucru este valabil indiferent dacă o celulă crește de una singură într-un iaz sau dacă este una dintre multele celule care formează un organism mai mare. Pentru a răspunde în mod corespunzător la stimulii externi, celulele au dezvoltat mecanisme complexe de comunicare, astfel încât să poată primi un mesaj, să transfere informația prin membrana plasmatică și apoi să producă modificări în interiorul celulei ca răspuns la mesaj.
În organismele multicelulare, celulele trimit și primesc în mod constant mesaje chimice pentru a coordona acțiunile altor organe, țesuturi și celule. Capacitatea de a trimite mesaje rapid și eficient permite celulelor să își coordoneze și să își ajusteze fin funcțiile. În timp ce necesitatea comunicării celulare în organismele mai mari pare evidentă, chiar și organismele unicelulare comunică între ele. Celulele de drojdie își transmit reciproc semnale pentru a ajuta la împerechere. Unele forme de bacterii își coordonează acțiunile pentru a forma complexe mari numite biofilme sau pentru a organiza producția de toxine pentru a elimina organismele concurente. Capacitatea celulelor de a comunica prin semnale chimice își are originea în celulele unice și a fost esențială pentru evoluția organismelor pluricelulare. Comunicarea eficientă, fără erori, este vitală pentru toate formele de viață.
- 9.1 | Molecule de semnalizare și receptori celulari
- 9.1.1 Forme de semnalizare
- Semnalizare paracrină
- Semnalizare autocrină
- Semnalizarea endocrină
- Semnalizare directă
- 9.1.2 Tipuri de receptori
- Receptorii interni
- Receptorii de suprafață celulară
- 9.1.3 Molecule de semnalizare
- Liganzii hidrofobi mici
- Ligani solubili în apă
- 9.2 | Propagarea semnalului
- 9.2.1 Căile de semnalizare și amplificarea semnalului
- 9.2.2 Metode de semnalizare intracelulară
- Fosforilarea
- Mesageri secundari
- 9.3 | Răspunsul la semnal
- 9.3.1 Răspunsuri la calea de semnalizare
- Expresia genică
- Creșterea metabolismului celular
- Creșterea celulară
- Moartea celulară
- 9.3.2 Încetarea căilor de semnalizare
9.1 | Molecule de semnalizare și receptori celulari
Până la sfârșitul acestei secțiuni, veți fi capabili să:
- Descris patru tipuri de semnalizare întâlnite în organismele multicelulare.
- Comparați receptorii interni cu receptorii de pe suprafața celulară.
- Recunoașteți relația dintre chimia unui ligand și mecanismul său de acțiune.
Există două tipuri de comunicare în lumea celulelor vii. Comunicarea între celule se numește semnalizare intercelulară, iar comunicarea în interiorul unei celule se numește semnalizare intracelulară. O modalitate ușoară de a reține distincția este înțelegerea faptului că prefixul inter- înseamnă „între” (o autostradă interstatală traversează mai multe state) și intra- înseamnă „înăuntru” (o perfuzie înseamnă intravenos sau „în interiorul venei”).
Semnale chimice sunt eliberate de o celulă de semnalizare și recepționate de o celulă țintă. Celulele țintă au proteine numite receptori, care se leagă de moleculele de semnalizare și provoacă un răspuns. Moleculele de semnalizare care se leagă de receptori se numesc liganzi. Liganzii și receptorii sunt specifici unul pentru celălalt; în mod obișnuit, un receptor se va lega numai la ligandul său specific. Cu toate acestea, există diferite tipuri de semnalizare.
9.1.1 Forme de semnalizare
Există patru categorii de semnalizare chimică întâlnite în organismele multicelulare: semnalizare autocrină, semnalizare paracrină, semnalizare endocrină și semnalizare directă prin joncțiunile gap (figura 9.2). Principala diferență între diferitele categorii de semnalizare este distanța pe care o parcurge semnalul pentru a ajunge la celula țintă.
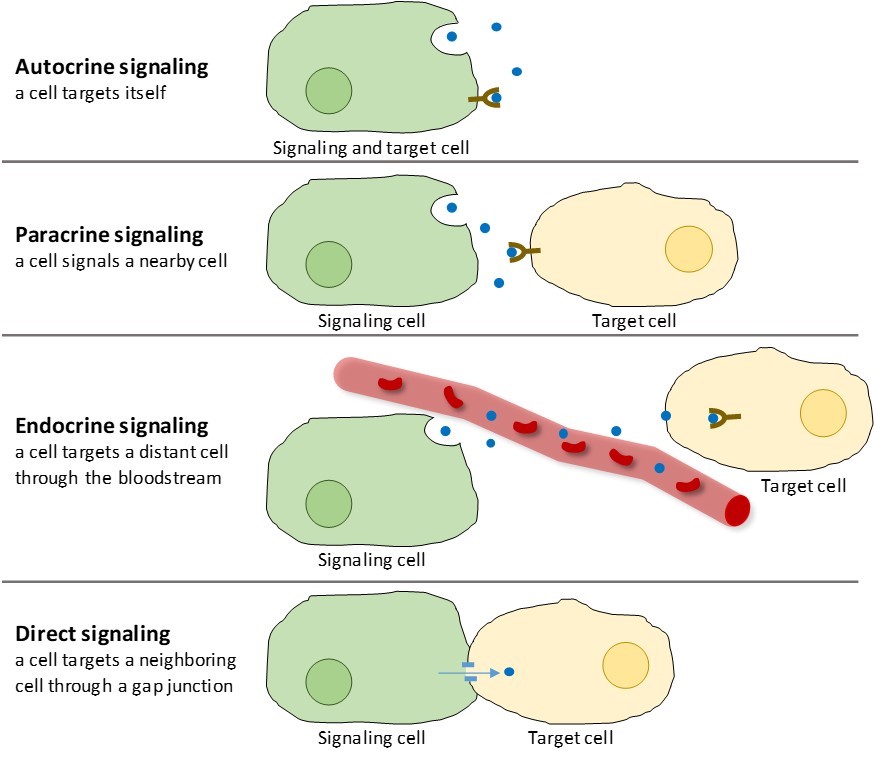
Semnalizare paracrină
Semnalurile care acționează local între celule care sunt apropiate se numesc semnale paracrine. Semnalele paracrine se deplasează prin difuzie prin matricea extracelulară (figura 9.2). Aceste tipuri de semnale determină, de obicei, răspunsuri rapide care durează doar o perioadă scurtă de timp. Pentru a menține răspunsul localizat, liganzii paracrini sunt de obicei degradați rapid de enzime sau eliminați de celulele vecine. Îndepărtarea semnalelor restabilește gradientul de concentrație pentru molecula semnal, permițându-le să difuzeze rapid prin spațiul intracelular dacă sunt eliberate din nou.
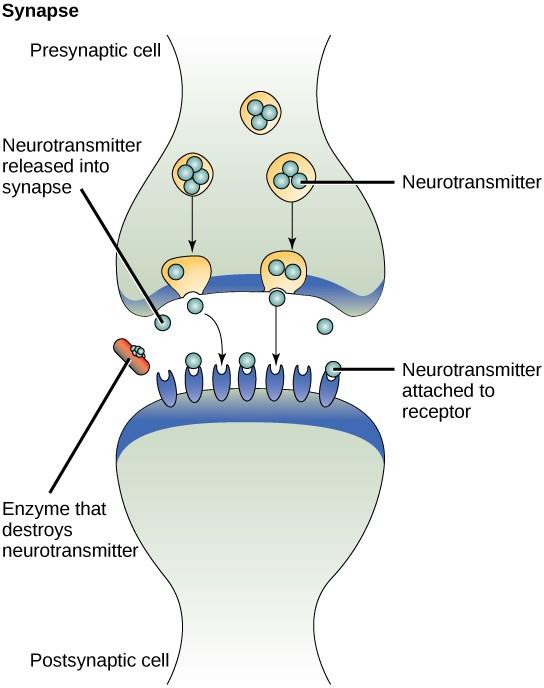
Un exemplu de semnalizare paracrină este transferul de semnale între celulele nervoase. Micul spațiu dintre celulele nervoase în care are loc transmiterea semnalelor se numește sinapsă. Semnalele sunt propagate de-a lungul celulelor nervoase prin impulsuri electrice cu mișcare rapidă. Atunci când aceste impulsuri ajung la capătul unei celule nervoase, liganzii chimici numiți neurotransmițători sunt eliberați în sinapsă de către celula presinaptică (celula care emite semnalul). Neurotransmițătorii se difuzează de-a lungul sinapsei (figura 9.3). Distanța mică dintre celulele nervoase permite semnalului să călătorească rapid, ceea ce permite un răspuns imediat, cum ar fi: „ia mâna de pe aragaz!”. Atunci când neurotransmițătorul se leagă de receptorul de pe suprafața celulei postsinaptice, este lansat următorul impuls electric. Neurotransmițătorii sunt degradați rapid sau sunt reabsorbiți de către celula presinaptică, astfel încât celula nervoasă receptoare să se recupereze rapid și să fie pregătită să răspundă rapid la următorul semnal sinaptic.
Semnalizare autocrină
Când o celulă răspunde la propria moleculă de semnalizare, aceasta se numește semnalizare autocrină (auto = „self”). Semnalizarea autocrină apare adesea împreună cu alte tipuri de semnalizare. De exemplu, atunci când este eliberat un semnal paracrin, celula de semnalizare poate răspunde la semnal împreună cu vecinii săi (figura 9.2).
Semnalizarea autocrină apare adesea în timpul dezvoltării timpurii a unui organism pentru a se asigura că celulele se dezvoltă în țesuturile corecte. Semnalizarea autocrină reglează, de asemenea, senzația de durere și răspunsurile inflamatorii. Mai mult, dacă o celulă este infectată cu un virus, celula se poate semnala pe sine pentru a fi supusă morții celulare programate, ucigând virusul în acest proces.
Semnalizarea endocrină
Semnalurile provenite de la celule îndepărtate se numesc semnale endocrine și provin din celulele endocrine. (În organism, multe celule endocrine sunt localizate în glandele endocrine, cum ar fi glanda tiroidă, hipotalamusul și glanda pituitară). Aceste tipuri de semnale produc de obicei un răspuns mai lent, dar au un efect de durată mai lungă. Liganzii eliberați în cadrul semnalizării endocrine se numesc hormoni, molecule de semnalizare care sunt produse într-o parte a corpului, dar care afectează alte regiuni ale corpului la o anumită distanță (figura 9.2).
Hormonii parcurg distanțele mari dintre celulele endocrine și celulele lor țintă prin intermediul fluxului sanguin, care este un mod relativ lent de a se deplasa în tot corpul. Din cauza formei lor de transport, hormonii se diluează și sunt prezenți în concentrații scăzute atunci când acționează asupra celulelor lor țintă. Acest lucru este diferit de semnalizarea paracrină, în care concentrațiile locale ale moleculelor de semnalizare pot fi foarte mari.
Semnalizare directă
Joncțiunile gap la animale și plasmodesmata la plante sunt conexiuni între membranele plasmatice ale celulelor vecine. Aceste canale umplute cu apă permit moleculelor mici de semnalizare să difuzeze între cele două celule. Moleculele mici, cum ar fi ionii de calciu (Ca2+), sunt capabile să se deplaseze între celule, dar moleculele mari, cum ar fi proteinele și ADN-ul, nu pot trece prin canale. Specificitatea canalelor asigură faptul că celulele rămân independente, dar pot transmite rapid și ușor semnale. Semnalarea directă permite unui grup de celule să își coordoneze răspunsul la un semnal pe care doar una dintre ele l-ar fi putut primi. La plante, plasmodesmele sunt omniprezente, transformând întreaga plantă într-o rețea gigantică de comunicare.
9.1.2 Tipuri de receptori
Receptorii sunt molecule proteice din celula țintă sau de pe suprafața acesteia care se leagă de liganzi. Există două tipuri de receptori, receptori interni și receptori de pe suprafața celulară.
Receptorii interni
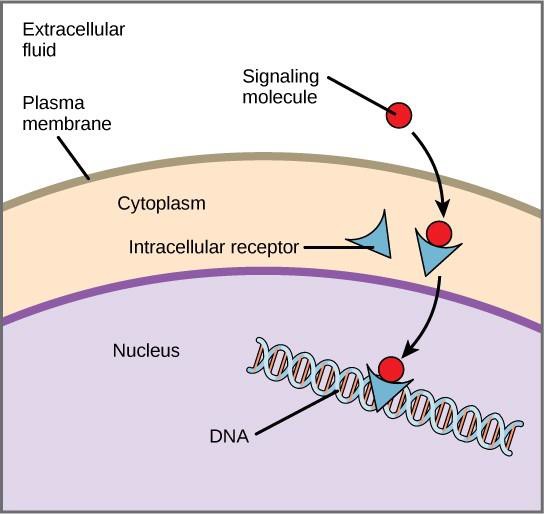
Receptorii interni, cunoscuți și sub numele de receptori intracelulari sau citoplasmatici, se găsesc în citoplasma celulelor țintă și răspund la moleculele de ligand hidrofobice care sunt capabile să traverseze membrana plasmatică. Odată intrate în interiorul celulei, multe dintre aceste molecule se leagă de proteine care acționează ca regulatori ai sintezei ARNm (transcripție) pentru a media expresia genelor.
Expresia genică este procesul celular de transformare a informațiilor din ADN-ul unei celule într-o secvență de aminoacizi, care formează în final o proteină. Atunci când ligandul se leagă de receptorul intern, se declanșează o modificare conformațională care expune un situs de legare la ADN pe proteina receptorului. Complexul ligand-receptor se deplasează în nucleu, apoi se leagă de regiuni de reglare specifice din ADN-ul cromozomial și promovează inițierea transcripției (figura 9.4). Transcripția este procesul de copiere a informațiilor din ADN-ul unei celule într-o formă specială de ARN numită ARN mesager (ARNm); celula utilizează informațiile din ARNm pentru a lega aminoacizii specifici în ordinea corectă, producând o proteină. Astfel, atunci când un ligand se leagă de un receptor intern, acesta poate influența direct expresia genetică în celula țintă.
Receptorii de suprafață celulară
Receptorii de suprafață celulară, cunoscuți și sub numele de receptori transmembranari, sunt proteine integrale care se leagă de moleculele de semnalizare externe. Acești receptori traversează membrana plasmatică și realizează transducția semnalului, în care un semnal extracelular este transformat într-un semnal intercelular. (Figura 9.5). Deoarece proteinele receptoare de la suprafața celulară sunt fundamentale pentru funcționarea normală a celulelor, nu ar trebui să fie o surpriză faptul că o funcționare defectuoasă a oricăreia dintre aceste proteine ar putea avea consecințe grave. S-a demonstrat că erorile în structurile proteice ale anumitor molecule receptoare joacă un rol în hipertensiune (tensiune arterială ridicată), astm, boli de inimă și cancer.
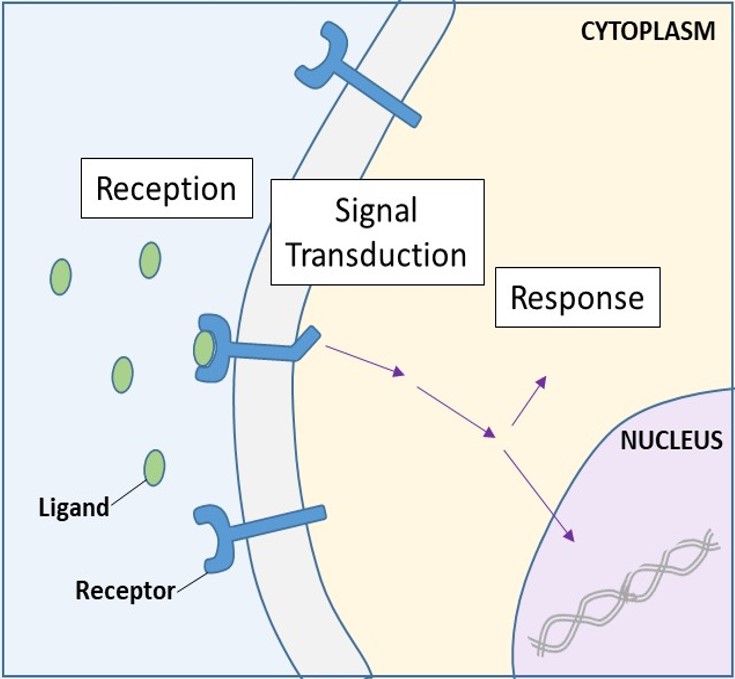
Care receptor de pe suprafața celulară are trei componente principale: un domeniu extern de legare a ligandului, sau domeniu extracelular; o regiune hidrofobă care traversează membrana; și un domeniu intracelular. Receptorii de pe suprafața celulară sunt implicați în cea mai mare parte a semnalizării în organismele multicelulare. Există trei categorii generale de receptori de suprafață celulară: receptori legați de enzime, receptori legați de canale ionice și receptori legați de proteinele G.
Receptorii legați de enzime sunt receptori de suprafață celulară cu domenii intracelulare care sunt asociați cu o enzimă. În unele cazuri, domeniul intracelular al receptorului însuși este o enzimă. Alți receptori legați de enzime au un mic domeniu intracelular care interacționează direct cu o enzimă. Receptorii legați de enzime au, în mod normal, domenii extracelulare și intracelulare mari, dar regiunea care traversează membrana este formată dintr-un singur alfa-helix în lanțul peptidic.
Când un ligand se leagă de domeniul extracelular al unui receptor legat de enzime, un semnal este transferat prin membrană, activând enzima. Activarea enzimei declanșează un lanț de evenimente în interiorul celulei care, în cele din urmă, conduce la un răspuns.
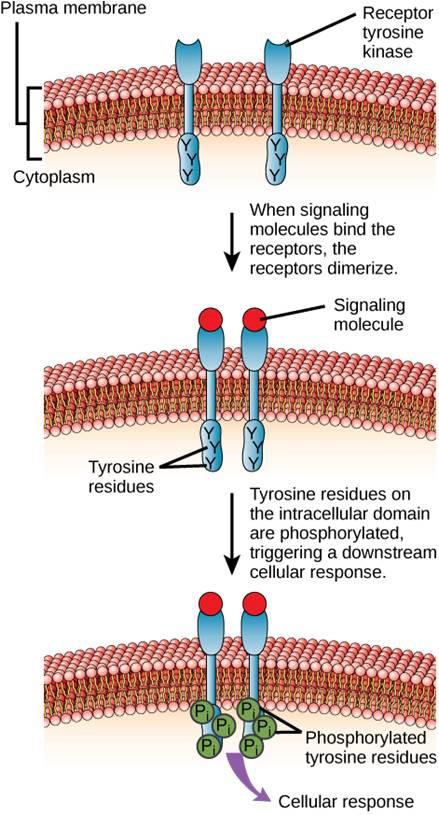
Un exemplu de receptor legat de o enzimă este receptorul tirosin kinazei (figura 9.6). O kinază este o enzimă care transferă grupările fosfat de la ATP la o altă proteină. Receptorul tirozin kinazei transferă grupări fosfat la moleculele de tirozină. Mai întâi, moleculele de semnalizare se leagă de domeniul extracelular a doi receptori de tirozin kinază aflați în apropiere. Cei doi receptori vecini se leagă apoi între ei sau se dimerizează. Fosfații sunt apoi adăugați la reziduurile de tirozină de pe domeniul intracelular al receptorilor (fosforilare). Reziduurile fosforilate pot apoi să transmită semnalul către următorul mesager din citoplasmă.
Receptorii factorului de creștere epidermică sunt un exemplu de receptori tirosin-kinaza care urmează acest mod de semnalizare. Defectele în semnalizarea ErbB din această familie pot duce la boli neuromusculare, cum ar fi scleroza multiplă și boala Alzheimer.
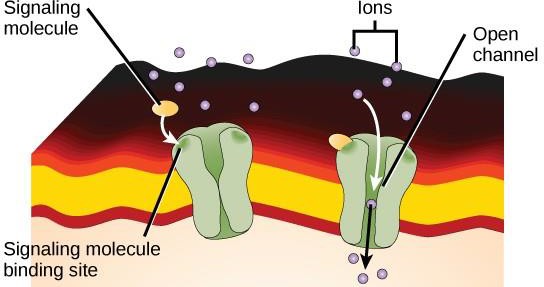
Receptorii legați de canale ionice se leagă de un ligand și deschid un canal prin membrană care permite trecerea unor ioni specifici. Acest tip de receptor de suprafață celulară are o regiune extinsă care se întinde pe membrană cu aminoacizi hidrofobi. În schimb, aminoacizii care căptușesc interiorul canalului sunt hidrofili pentru a permite trecerea ionilor. Atunci când un ligand se leagă de regiunea extracelulară a canalului, are loc o modificare conformațională în structura proteinei care permite trecerea ionilor, cum ar fi sodiul, calciul, magneziul sau hidrogenul (figura 9.7).
Receptorii legați de proteina G se leagă de un ligand și activează o proteină G asociată. Proteina G activată interacționează apoi cu o proteină membranară din apropiere, care poate fi un canal ionic sau o enzimă (figura 9.8). Toți receptorii legați de proteina G au șapte domenii transmembranare, dar fiecare receptor are un domeniu extracelular specific și un situs de legare a proteinei G.
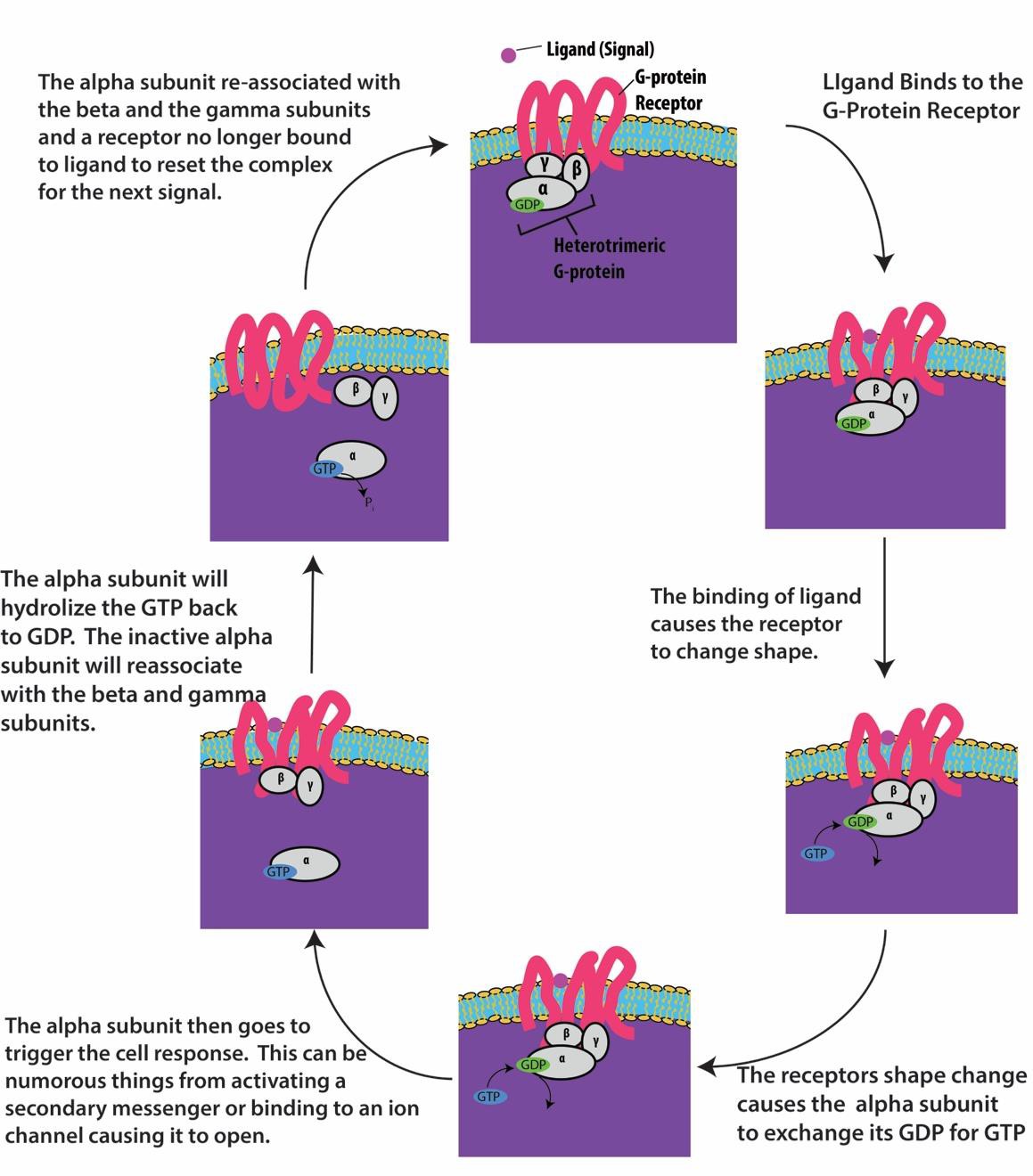
Semnalizarea celulară cu ajutorul receptorilor legați de proteina G are loc sub forma unui ciclu. Odată ce ligandul se leagă de receptor, schimbarea de formă rezultată activează proteina G, care eliberează GDP și preia GTP. Subunitățile proteinei G se împart apoi în subunitățile α și βγ. Unul sau ambele fragmente ale acestei proteine G pot fi capabile să activeze alte proteine din celulă. După un timp, GTP-ul de pe subunitatea α activă a proteinei G este hidrolizat în GDP, iar subunitatea βγ este dezactivată. Subunitățile se reasociază pentru a forma proteina G inactivă și ciclul începe din nou (figura 9.8).
Receptorii legați de proteina G sunt utilizați în multe procese fiziologice, inclusiv în cele de transducție a vederii, a gustului și de reglare a sistemului imunitar și a inflamației.

HER2 este un receptor tirozin-kinază. În 30 la sută dintre cancerele mamare umane, HER2 este activat permanent, ceea ce duce la o diviziune celulară nereglementată. Lapatinib, un medicament utilizat pentru tratarea cancerului de sân, inhibă procesul prin care receptorul se fosforilează, reducând astfel creșterea tumorii cu 50 la sută. Care dintre următoarele etape ar fi inhibate de Lapatinib?
- Legatura moleculei de semnalizare, dimerizarea și răspunsul celular din aval
- Dimerizarea și răspunsul celular din aval
- Răspunsul celular din aval
- Activitatea fosfatazei, dimerizarea și răspunsul celular din aval
9.1.3 Molecule de semnalizare
Produse de celulele de semnalizare, liganzii sunt semnale chimice care se deplasează către celulele țintă și provoacă un răspuns. Tipurile de molecule care servesc drept liganzi sunt incredibil de variate și variază de la proteine mici la ioni mici. Liganzii sunt clasificați fie ca liganzi hidrofobi mici, care pot traversa membranele plasmatice, fie ca liganzi solubili în apă, care nu pot.
Liganzii hidrofobi mici
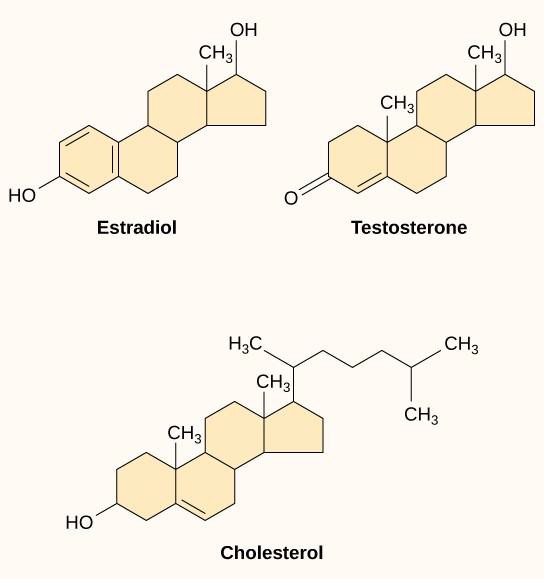
Liganzii hidrofobi mici, numiți și liganzi solubili în lipide, pot difuza direct prin membrana plasmatică și interacționa cu receptorii interni. Membri importanți ai acestei clase de liganzi sunt hormonii steroizi. Steroizii sunt lipide care au un schelet hidrocarbonat cu patru inele fuzionate; diferiți steroizi au diferite grupe funcționale atașate la scheletul de carbon. Hormonii steroizi includ hormonul sexual feminin estradiolul, care este un tip de estrogen; hormonul sexual masculin testosteronul; și colesterolul, care este o componentă structurală importantă a membranelor biologice și un precursor al hormonilor steroizi (figura 9.10). Alți hormoni hidrofobi includ hormonii tiroidieni și vitamina D. Pentru a fi solubili în sânge, liganzii hidrofobi trebuie să se lege de proteinele purtătoare în timp ce sunt transportați prin fluxul sanguin.
Ligani solubili în apă
Din moment ce liganzii solubili în apă sunt polari, ei nu pot trece prin membrana plasmatică fără ajutor. Uneori sunt prea mari pentru a trece deloc prin membrană. În schimb, majoritatea liganzilor hidrosolubili se leagă de domeniul extracelular al receptorilor de pe suprafața celulară (a se vedea figura 9.5). Acest grup de liganzi este destul de divers și include molecule mici, peptide și proteine.
9.2 | Propagarea semnalului
Până la sfârșitul acestei secțiuni, veți fi capabili să:
- Explicați modul în care legarea unui ligand inițiază transducția semnalului în întreaga celulă.
- Recunoașteți că semnalele intracelulare sunt transmise prin rolul fosforilării sau al unor mesageri secundari.
După ce un ligand hidrosolubil se leagă de receptorul său, semnalul este transmis prin membrană și în citoplasmă. Continuarea unui semnal în acest mod se numește transducție de semnal (figura 9.5). Transducția semnalului are loc numai cu receptorii de la suprafața celulară, deoarece receptorii interni sunt capabili să pătrundă în celulă.
Când un ligand se leagă de receptorul său, au loc modificări conformaționale care afectează domeniul intracelular al receptorului. Aceste modificări conformaționale conduc la activarea domeniului intracelular sau a proteinelor asociate acestuia. În unele cazuri, legarea ligandului determină dimerizarea receptorului, ceea ce înseamnă că doi receptori se leagă unul de celălalt pentru a forma un complex stabil numit dimer. Un dimer este un compus chimic format atunci când două molecule (adesea identice) se unesc. Legarea receptorilor în acest mod permite domeniilor lor intracelulare să intre în contact strâns și să se activeze reciproc.
9.2.1 Căile de semnalizare și amplificarea semnalului
Deși moleculele de semnalizare se găsesc adesea la concentrații foarte mici, ele pot produce efecte profunde. După ce ligandul se leagă de receptorul de pe suprafața celulară, activarea componentelor intracelulare ale receptorului declanșează un lanț de evenimente care se numește cale de semnalizare sau cascadă de semnalizare. Într-o cale de semnalizare, mesagerii secundari, enzimele și/sau proteinele activate activează alte proteine sau mesageri (figura 9.11). Fiecare membru al căii poate activa mii de membri ai următorului membru al căii, într-un proces numit amplificare a semnalului. Deoarece semnalul este amplificat la fiecare etapă, se poate genera un răspuns foarte mare de la un singur receptor care leagă un ligand.
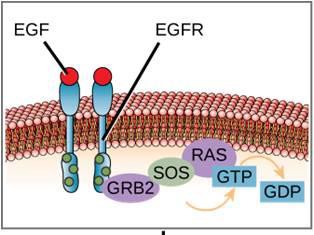
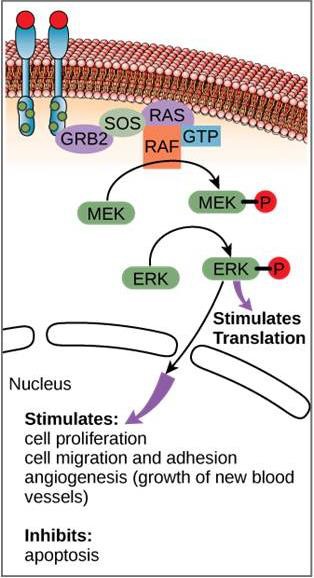
Un exemplu de cale de semnalizare este prezentat în figura 9.11. Factorul de creștere epidermică (EGF) este o moleculă de semnalizare care este implicată în reglarea creșterii celulare, vindecarea rănilor și repararea țesuturilor. Receptorul pentru EGF (EGFR) este o tirosin-kinază. O kinază activată fosforilează și activează multe molecule din aval. Atunci când EGF se leagă de EGFR, o cascadă de evenimente de fosforilare în aval semnalează celulei să crească și să se divizeze. Dacă EGFR este activat în momente nepotrivite, se poate produce o creștere necontrolată a celulelor (cancer).
În anumite tipuri de cancer, activitatea GTPază a proteinei RAS G este inhibată. Aceasta înseamnă că proteina RAS nu mai poate hidroliza GTP în GDP. Ce efect ar avea acest lucru asupra evenimentelor celulare din aval?
9.2.2 Metode de semnalizare intracelulară
Inducția unei căi de semnalizare depinde de modificarea unei componente celulare de către o enzimă. Există numeroase modificări enzimatice care pot avea loc pentru a activa următoarea componentă a căii. Următoarele sunt unele dintre cele mai frecvente evenimente din cadrul semnalizării intracelulare.
Fosforilarea
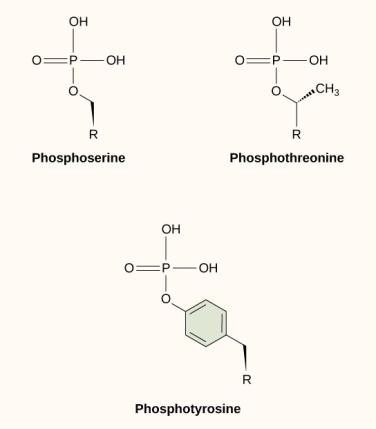
Una dintre cele mai frecvente modificări chimice care are loc în căile de semnalizare este adăugarea unei grupări fosfat la o moleculă într-un proces numit fosforilare. Fosfatul poate fi adăugat la o nucleotidă, cum ar fi GMP, pentru a forma GDP sau GTP. De asemenea, fosfații sunt adesea adăugați la reziduurile de serină, treonină și tirozină din proteine, unde înlocuiesc gruparea hidroxil a aminoacidului (figura 9.12). Transferul fosfatului este catalizat de o enzimă numită kinază. Fosforilarea poate activa sau inactiva enzimele, iar inversarea fosforilării, defosforilarea, va inversa efectul.
Mesageri secundari
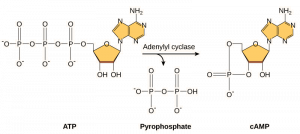
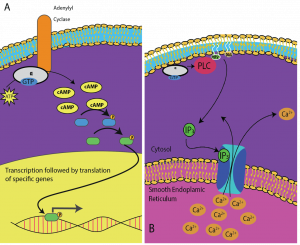
Mesagerii secundari sunt molecule mici care propagă un semnal după ce acesta a fost inițiat prin legarea moleculei de semnalizare la receptor. Aceste molecule ajută la propagarea unui semnal prin citoplasmă prin modificarea comportamentului anumitor proteine celulare. Un al doilea mesager utilizat de multe tipuri de celule diferite este AMP ciclic (AMPc). AMP ciclic este sintetizat de către enzima adenililciclază din ATP (figura 9.13). Rolul principal al AMPc în celule este acela de a se lega și de a activa o enzimă numită kinază dependentă de AMPc (A-kinază). A-kinaza reglează multe căi metabolice vitale: Ea fosforilează reziduurile de serină și treonină ale proteinelor sale țintă, activându-le în acest proces. A-kinaza se găsește în multe tipuri diferite de celule, iar proteinele țintă din fiecare tip de celulă sunt diferite. Un alt mesager secundar este Ca2+care poate fi eliberat pentru a inunda celula.
Celule diferite răspund diferit la AMPc. În figura 9.14, este prezentată subunitatea alfa de la un receptor al proteinei G care activează două tipuri diferite de semnalizare. În prima imagine, AMPc este produs de enzima adenilat ciclază atunci când este activată de subunitatea alfa. AMPc activează apoi alte proteine care afectează transcrierea genelor. În cea de-a doua imagine, subunitatea alfa de la proteina G declanșează o cascadă care eliberează Ca2+ din reticulul endoplasmatic neted. În acest caz, Ca2+ este mesagerul secundar care determină răspunsul celular.
9.3 | Răspunsul la semnal
Până la sfârșitul acestei secțiuni, veți fi capabili să:
- Recunoașteți că căile de semnalizare dirijează expresia proteinelor, metabolismul celular și creșterea celulară.
- Recunoașteți rolul apoptozei în dezvoltarea și menținerea unui organism sănătos.
Utilizând căile de transducție a semnalelor, receptorii din membrana plasmatică produc o varietate de efecte asupra celulei. În interiorul celulei, liganzii se leagă de receptorii lor interni, permițându-le să afecteze direct ADN-ul celulei și mașinăria de producere a proteinelor. Rezultatele căilor de semnalizare sunt extrem de variate și depind de tipul de celulă implicat, precum și de condițiile externe și interne. Un mic eșantion de răspunsuri este descris mai jos.
9.3.1 Răspunsuri la calea de semnalizare
Expresia genică
Câteva căi de transducție a semnalului reglează transcrierea ARN-ului. Altele reglează traducerea proteinelor.
Creșterea metabolismului celular
Activarea receptorilor β-adrenergici din celulele musculare de către adrenalină duce la o creștere a AMP ciclic în interiorul celulei. Adrenalina este un hormon produs de glanda suprarenală care pregătește organismul pentru urgențe pe termen scurt.
Creșterea celulară
Căile de semnalizare celulară joacă, de asemenea, un rol major în diviziunea celulară. Celulele nu se divid în mod normal decât dacă sunt stimulate de semnale de la alte celule. Liganzii care promovează creșterea celulară se numesc factori de creștere. Majoritatea factorilor de creștere se leagă de receptori de pe suprafața celulară care sunt legați de tirozin-kinaze.
Moartea celulară
Când o celulă este deteriorată, superfluă sau potențial periculoasă pentru un organism, o celulă poate iniția un mecanism pentru a declanșa moartea celulară programată sau apoptoza. Apoptoza permite ca o celulă să moară într-un mod controlat care previne eliberarea de molecule potențial dăunătoare din interiorul celulei. Cu toate acestea, în unele cazuri, cum ar fi o infecție virală sau o diviziune necontrolată a celulelor din cauza cancerului, controalele și echilibrele normale ale celulei eșuează. Semnalizarea externă poate, de asemenea, să inițieze apoptoza. De exemplu, majoritatea celulelor animale normale au receptori care interacționează cu matricea extracelulară, o rețea de glicoproteine care asigură suportul structural pentru celulele animale. Legarea receptorilor celulari de matricea extracelulară inițiază o cascadă de semnalizare în interiorul celulei. Cu toate acestea, dacă celula se îndepărtează de matricea extracelulară, semnalizarea încetează, iar celula suferă apoptoză. Acest sistem ajută la prevenirea deplasării celulelor prin organism și a proliferării scăpate de sub control, așa cum se întâmplă în cazul celulelor tumorale care fac metastaze.
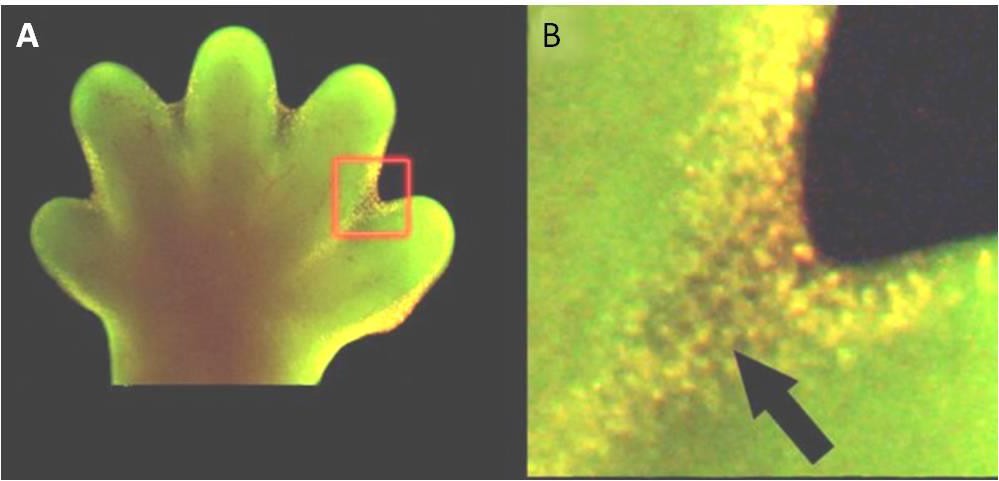
Apoptoza este, de asemenea, esențială pentru dezvoltarea embriologică normală. La vertebrate, de exemplu, stadiile timpurii de dezvoltare includ formarea țesutului asemănător unei pânze între degetele individuale de la mâini și picioare (figura 9.15). Pe parcursul dezvoltării normale, aceste celule inutile trebuie să fie eliminate, permițând formarea degetelor de la mâini și picioare complet separate. Un mecanism de semnalizare celulară declanșează apoptoza, care distruge celulele dintre degetele în curs de dezvoltare.
9.3.2 Încetarea căilor de semnalizare
Semnalizarea aberantă observată adesea în celulele tumorale este o dovadă că încetarea unui semnal la momentul potrivit poate fi la fel de importantă ca și inițierea unui semnal. O metodă de a opri un semnal specific este de a degrada ligandul sau de a-l elimina astfel încât să nu mai poată accesa receptorul său. Unul dintre motivele pentru care hormonii hidrofobi, cum ar fi estrogenul și testosteronul, declanșează evenimente de lungă durată este faptul că se leagă de proteinele purtătoare. Aceste proteine permit moleculelor insolubile să fie solubile în sânge, dar ele protejează, de asemenea, hormonii de degradarea de către enzimele circulante.
În interiorul celulei, multe enzime diferite inversează modificările celulare care rezultă din cascadele de semnalizare. De exemplu, fosfatazele sunt enzime care îndepărtează gruparea fosfat atașată proteinelor de către kinaze într-un proces numit defosforilare. cAMP este degradat în AMP de către fosfodiesterază, punând astfel capăt semnalului său. În mod similar, eliberarea depozitelor de calciu este inversată de pompele de Ca2+ care sunt localizate în membranele celulare.
.