
Scrivi qui i tuoi esempi.
- 9.1 Molecole di segnalazione e recettori cellulari
- 9.2 Propagazione del segnale
- 9.3 Risposta al segnale
Immagina come sarebbe la vita se tu e le persone intorno a te non poteste comunicare. Non sareste in grado di esprimere i vostri desideri agli altri, né potreste fare domande per saperne di più sul vostro ambiente. L’organizzazione sociale dipende dalla comunicazione tra gli individui che compongono quella società.
Come per le persone, è vitale per le singole cellule essere in grado di interagire con il loro ambiente e tra di loro. Questo è vero sia che una cellula cresca da sola in uno stagno sia che sia una delle tante cellule che formano un organismo più grande. Per rispondere correttamente agli stimoli esterni, le cellule hanno sviluppato complessi meccanismi di comunicazione in modo da poter ricevere un messaggio, trasferire le informazioni attraverso la membrana plasmatica e poi produrre cambiamenti all’interno della cellula in risposta al messaggio.
Negli organismi multicellulari, le cellule inviano e ricevono costantemente messaggi chimici per coordinare le azioni di altri organi, tessuti e cellule. La capacità di inviare messaggi in modo rapido ed efficiente permette alle cellule di coordinare e mettere a punto le loro funzioni. Mentre la necessità della comunicazione cellulare negli organismi più grandi sembra ovvia, anche gli organismi unicellulari comunicano tra loro. Le cellule di lievito si segnalano a vicenda per favorire l’accoppiamento. Alcune forme di batteri coordinano le loro azioni per formare grandi complessi chiamati biofilm o per organizzare la produzione di tossine per rimuovere gli organismi concorrenti. La capacità delle cellule di comunicare attraverso segnali chimici ha avuto origine nelle cellule singole ed è stata essenziale per l’evoluzione degli organismi multicellulari. Una comunicazione efficiente e senza errori è vitale per tutta la vita.
- 9.1 | Molecole di segnalazione e recettori cellulari
- 9.1.1 Forme di segnalazione
- Segnali paracrini
- Segnalazione autocrina
- Segnalazioni endocrine
- Segnalazione diretta
- 9.1.2 Tipi di recettori
- Ricettori interni
- Ricettori di superficie cellulare
- 9.1.3 Molecole di segnalazione
- Ligandi idrofobici piccoli
- Ligandi solubili in acqua
- 9.2 | Propagazione del segnale
- 9.2.1 Vie di segnalazione e amplificazione del segnale
- 9.2.2 Metodi di segnalazione intracellulare
- Fosforilazione
- Secondi messaggeri
- 9.3 | Risposta al segnale
- 9.3.1 Risposte alla via di segnalazione
- Espressione genica
- Aumento del metabolismo cellulare
- Crescita cellulare
- Morte cellulare
- 9.3.2 Cessazione delle vie di segnalazione
9.1 | Molecole di segnalazione e recettori cellulari
Alla fine di questa sezione, sarai in grado di:
- Descrivere quattro tipi di segnalazione trovati negli organismi multicellulari.
- Confrontare i recettori interni con quelli della superficie cellulare.
- Riconoscere la relazione tra la chimica di un ligando e il suo meccanismo d’azione.
Ci sono due tipi di comunicazione nel mondo delle cellule viventi. La comunicazione tra le cellule è chiamata segnalazione intercellulare, e la comunicazione all’interno di una cellula è chiamata segnalazione intracellulare. Un modo semplice per ricordare la distinzione è capire che il prefisso inter- significa “tra” (un’autostrada interstatale attraversa diversi stati) e intra- significa “dentro” (una flebo significa intravenosa o “dentro la vena”).
I segnali chimici sono rilasciati da una cellula di segnalazione e ricevuti da una cellula bersaglio. Le cellule bersaglio hanno proteine chiamate recettori, che si legano alle molecole di segnalazione e causano una risposta. Le molecole di segnalazione che si legano ai recettori sono chiamate ligandi. I ligandi e i recettori sono specifici l’uno per l’altro; un recettore in genere si lega solo al suo ligando specifico. Tuttavia, ci sono diversi tipi di segnalazione.
9.1.1 Forme di segnalazione
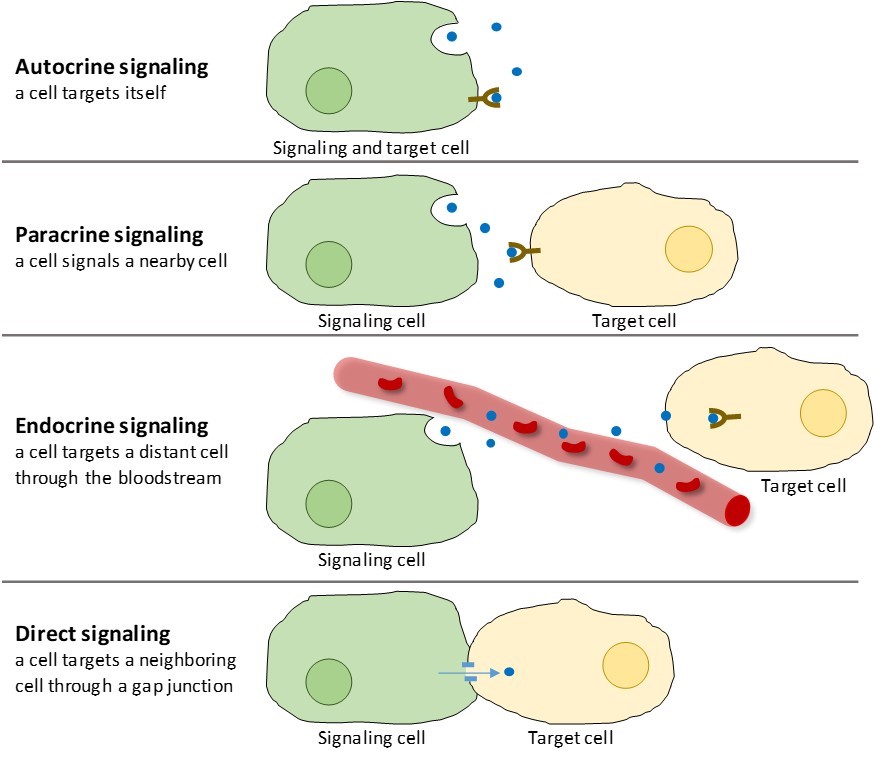
Ci sono quattro categorie di segnalazione chimica che si trovano negli organismi multicellulari: segnalazione autocrina, segnalazione paracrina, segnalazione endocrina e segnalazione diretta attraverso le giunzioni gap (Figura 9.2). La principale differenza tra le diverse categorie di segnalazione è la distanza che il segnale percorre per raggiungere la cellula bersaglio.
Segnali paracrini
I segnali che agiscono localmente tra cellule vicine sono chiamati segnali paracrini. I segnali paracrini si muovono per diffusione attraverso la matrice extracellulare (Figura 9.2). Questi tipi di segnali di solito suscitano risposte rapide che durano solo un breve periodo di tempo. Per mantenere la risposta localizzata, i ligandi paracrini vengono solitamente degradati rapidamente da enzimi o rimossi dalle cellule vicine. La rimozione dei segnali ristabilisce il gradiente di concentrazione per la molecola segnale, permettendole di diffondersi rapidamente attraverso lo spazio intracellulare se rilasciata nuovamente.
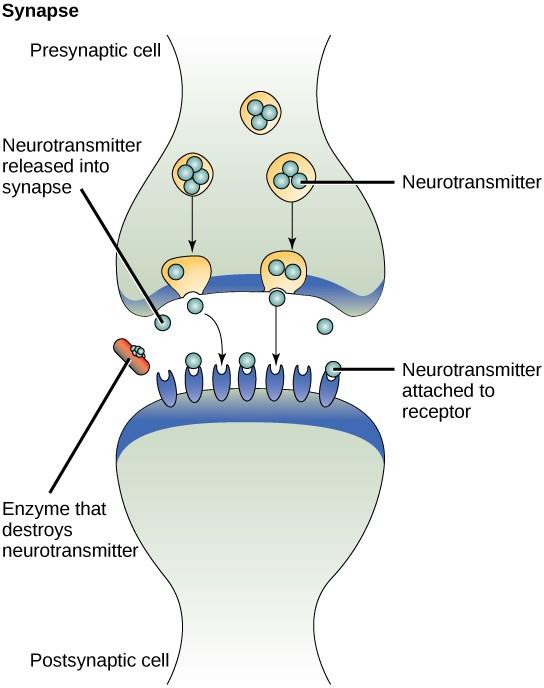
Un esempio di segnalazione paracrina è il trasferimento di segnali tra cellule nervose. Il minuscolo spazio tra le cellule nervose dove avviene la trasmissione del segnale è chiamato sinapsi. I segnali sono propagati lungo le cellule nervose da impulsi elettrici che si muovono velocemente. Quando questi impulsi raggiungono l’estremità di una cellula nervosa, ligandi chimici chiamati neurotrasmettitori vengono rilasciati nella sinapsi dalla cellula presinaptica (la cellula che emette il segnale). I neurotrasmettitori si diffondono attraverso la sinapsi (Figura 9.3). La piccola distanza tra le cellule nervose permette al segnale di viaggiare rapidamente, il che permette una risposta immediata, come “togli la mano dalla stufa!”. Quando il neurotrasmettitore lega il recettore sulla superficie della cellula postsinaptica, viene lanciato il successivo impulso elettrico. I neurotrasmettitori sono degradati rapidamente o sono riassorbiti dalla cellula presinaptica in modo che la cellula nervosa ricevente possa recuperare rapidamente ed essere pronta a rispondere rapidamente al prossimo segnale sinaptico.
Segnalazione autocrina
Quando una cellula risponde alla propria molecola di segnalazione, si chiama segnalazione autocrina (auto = “sé”). La segnalazione autocrina si verifica spesso con altri tipi di segnalazione. Per esempio, quando viene rilasciato un segnale paracrino, la cellula di segnalazione può rispondere al segnale insieme ai suoi vicini (Figura 9.2).
La segnalazione autocrina avviene spesso durante lo sviluppo iniziale di un organismo per assicurare che le cellule si sviluppino nei tessuti corretti. La segnalazione autocrina regola anche la sensazione di dolore e le risposte infiammatorie. Inoltre, se una cellula è infettata da un virus, la cellula può segnalare a se stessa di subire la morte cellulare programmata, uccidendo il virus nel processo.
Segnalazioni endocrine
I segnali da cellule lontane sono chiamati segnali endocrini, e hanno origine dalle cellule endocrine. (Nel corpo, molte cellule endocrine si trovano nelle ghiandole endocrine, come la tiroide, l’ipotalamo e l’ipofisi). Questi tipi di segnali di solito producono una risposta più lenta ma hanno un effetto più duraturo. I ligandi rilasciati nella segnalazione endocrina sono chiamati ormoni, molecole di segnalazione che sono prodotte in una parte del corpo ma influenzano altre regioni del corpo ad una certa distanza (Figura 9.2).
Gli ormoni percorrono le grandi distanze tra le cellule endocrine e le loro cellule bersaglio attraverso il flusso sanguigno, che è un modo relativamente lento per muoversi in tutto il corpo. A causa della loro forma di trasporto, gli ormoni si diluiscono e sono presenti in basse concentrazioni quando agiscono sulle loro cellule bersaglio. Questo è diverso dalla segnalazione paracrina, in cui le concentrazioni locali di molecole di segnalazione possono essere molto alte.
Segnalazione diretta
Le giunzioni di gap negli animali e i plasmodesmata nelle piante sono collegamenti tra le membrane plasmatiche di cellule vicine. Questi canali pieni d’acqua permettono a piccole molecole di segnalazione di diffondersi tra le due cellule. Le piccole molecole, come gli ioni di calcio (Ca2+), sono in grado di muoversi tra le cellule, ma le grandi molecole come le proteine e il DNA non possono passare attraverso i canali. La specificità dei canali assicura che le cellule rimangano indipendenti ma possano trasmettere segnali in modo rapido e semplice. La segnalazione diretta permette a un gruppo di cellule di coordinare la loro risposta a un segnale che solo una di loro può aver ricevuto. Nelle piante, i plasmodesmata sono ubiquitari, rendendo l’intera pianta una gigantesca rete di comunicazione.
9.1.2 Tipi di recettori
I recettori sono molecole proteiche nella cellula bersaglio o sulla sua superficie che si legano ai ligandi. Ci sono due tipi di recettori, quelli interni e quelli sulla superficie cellulare.
Ricettori interni
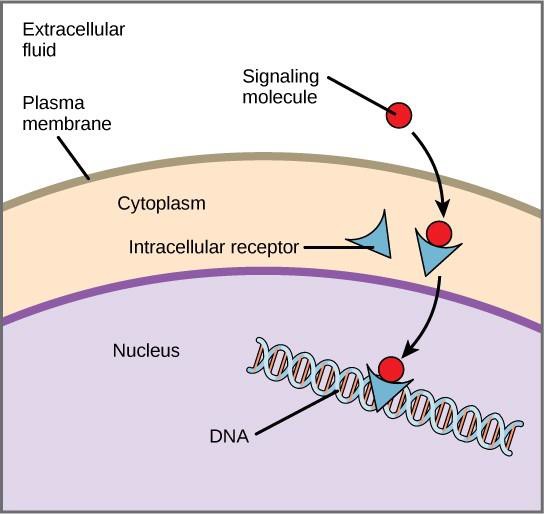
I recettori interni, conosciuti anche come recettori intracellulari o citoplasmatici, si trovano nel citoplasma delle cellule bersaglio e rispondono a molecole di ligandi idrofobici che sono in grado di viaggiare attraverso la membrana plasmatica. Una volta all’interno della cellula, molte di queste molecole si legano a proteine che agiscono come regolatori della sintesi dell’mRNA (trascrizione) per mediare l’espressione genica.
L’espressione genica è il processo cellulare di trasformazione delle informazioni nel DNA di una cellula in una sequenza di aminoacidi, che alla fine forma una proteina. Quando il ligando si lega al recettore interno, si innesca un cambiamento conformazionale che espone un sito di legame al DNA sulla proteina recettore. Il complesso ligando-recettore si sposta nel nucleo, poi si lega a specifiche regioni regolatrici del DNA cromosomico e promuove l’inizio della trascrizione (Figura 9.4). La trascrizione è il processo di copiatura delle informazioni nel DNA di una cellula in una forma speciale di RNA chiamata RNA messaggero (mRNA); la cellula usa le informazioni nell’mRNA per collegare specifici aminoacidi nell’ordine corretto, producendo una proteina. Così, quando un ligando si lega a un recettore interno, può influenzare direttamente l’espressione genica nella cellula bersaglio.
Ricettori di superficie cellulare
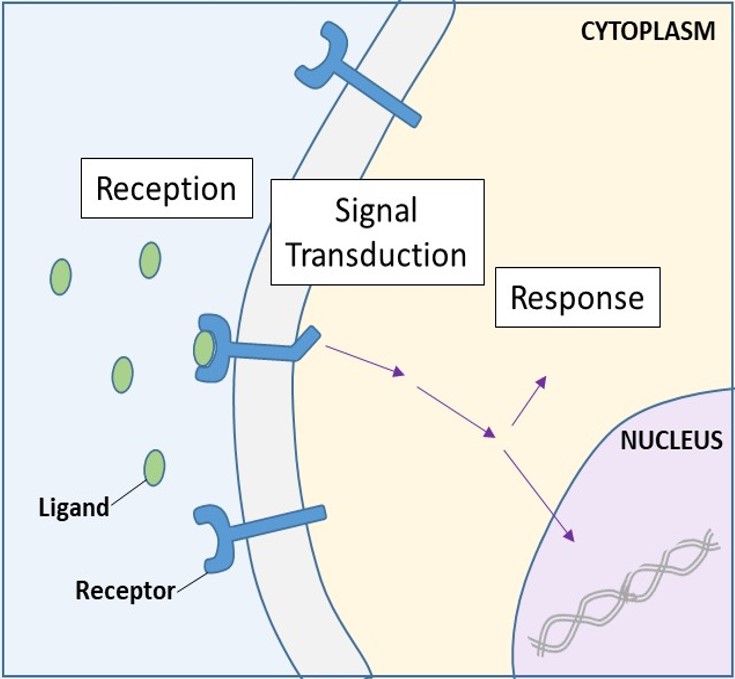
I recettori di superficie cellulare, noti anche come recettori transmembrana, sono proteine integrali che si legano a molecole di segnalazione esterne. Questi recettori attraversano la membrana plasmatica ed eseguono la trasduzione del segnale, in cui un segnale extracellulare viene convertito in un segnale intercellulare. (Figura 9.5). Poiché le proteine recettoriali della superficie cellulare sono fondamentali per il normale funzionamento delle cellule, non dovrebbe sorprendere che un malfunzionamento di una qualsiasi di queste proteine possa avere gravi conseguenze. È stato dimostrato che errori nella struttura proteica di alcune molecole recettoriali hanno un ruolo nell’ipertensione (pressione alta), nell’asma, nelle malattie cardiache e nel cancro.
Ogni recettore a superficie cellulare ha tre componenti principali: un dominio esterno che lega il ligando, o dominio extracellulare; una regione idrofoba che attraversa la membrana; e un dominio intracellulare. I recettori a superficie cellulare sono coinvolti nella maggior parte della segnalazione negli organismi multicellulari. Ci sono tre categorie generali di recettori a superficie cellulare: recettori legati all’enzima, recettori legati ai canali ionici e recettori legati alle proteine G.
I recettori legati all’enzima sono recettori a superficie cellulare con domini intracellulari che sono associati a un enzima. In alcuni casi, il dominio intracellulare del recettore stesso è un enzima. Altri recettori legati a un enzima hanno un piccolo dominio intracellulare che interagisce direttamente con un enzima. I recettori legati all’enzima hanno normalmente grandi domini extracellulari e intracellulari, ma la regione che attraversa la membrana consiste in una singola alfa-elica nel filamento peptidico.
Quando un ligando si lega al dominio extracellulare di un recettore legato all’enzima, un segnale viene trasferito attraverso la membrana, attivando l’enzima. L’attivazione dell’enzima innesca una catena di eventi all’interno della cellula che alla fine porta a una risposta.
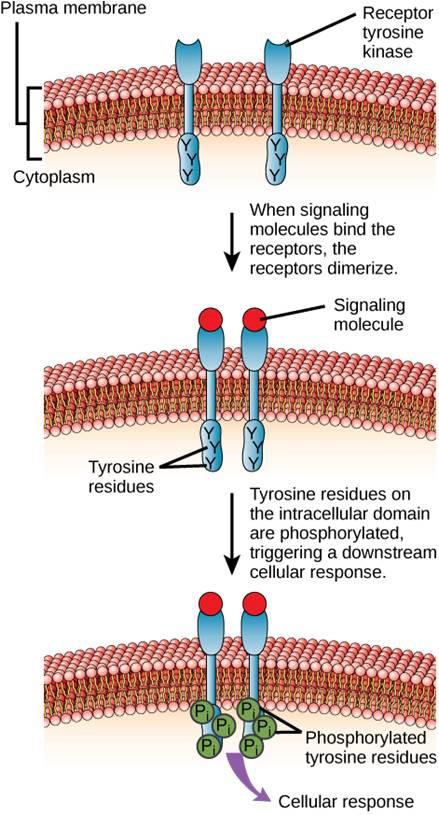
Un esempio di recettore legato a un enzima è il recettore tirosin-chinasi (Figura 9.6). Una chinasi è un enzima che trasferisce gruppi fosfato dall’ATP a un’altra proteina. Il recettore della tirosina chinasi trasferisce gruppi fosfato alle molecole di tirosina. In primo luogo, le molecole di segnalazione si legano al dominio extracellulare di due recettori tirosin-chinasi vicini. I due recettori vicini si legano poi insieme, o dimerizzano. I fosfati vengono poi aggiunti ai residui di tirosina sul dominio intracellulare dei recettori (fosforilazione). I residui fosforilati possono quindi trasmettere il segnale al prossimo messaggero all’interno del citoplasma.
I recettori del fattore di crescita epidermico sono un esempio di recettore tirosin-chinasi che segue questa modalità di segnalazione. Difetti nella segnalazione di ErbB in questa famiglia possono portare a malattie neuromuscolari come la sclerosi multipla e il morbo di Alzheimer.
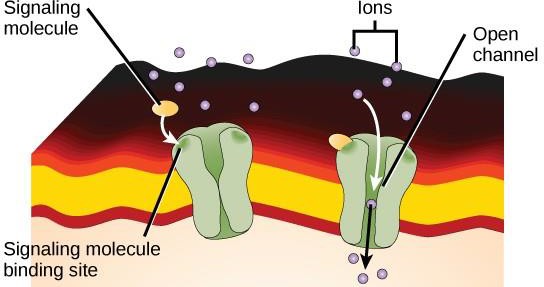
I recettori legati a canali ionici si legano a un ligando e aprono un canale attraverso la membrana che permette il passaggio di ioni specifici. Questo tipo di recettore a superficie cellulare ha un’ampia regione che attraversa la membrana con aminoacidi idrofobici. Al contrario, gli amminoacidi che rivestono l’interno del canale sono idrofili per permettere il passaggio degli ioni. Quando un ligando si lega alla regione extracellulare del canale, si verifica un cambiamento conformazionale nella struttura della proteina che permette il passaggio di ioni come il sodio, il calcio, il magnesio o l’idrogeno (Figura 9.7).
I recettori legati alle proteine G si legano a un ligando e attivano una proteina G associata. La proteina G attivata interagisce poi con una proteina di membrana vicina, che può essere un canale ionico o un enzima (Figura 9.8). Tutti i recettori legati alle proteine G hanno sette domini transmembrana, ma ogni recettore ha un dominio extracellulare specifico e un sito di legame alle proteine G.
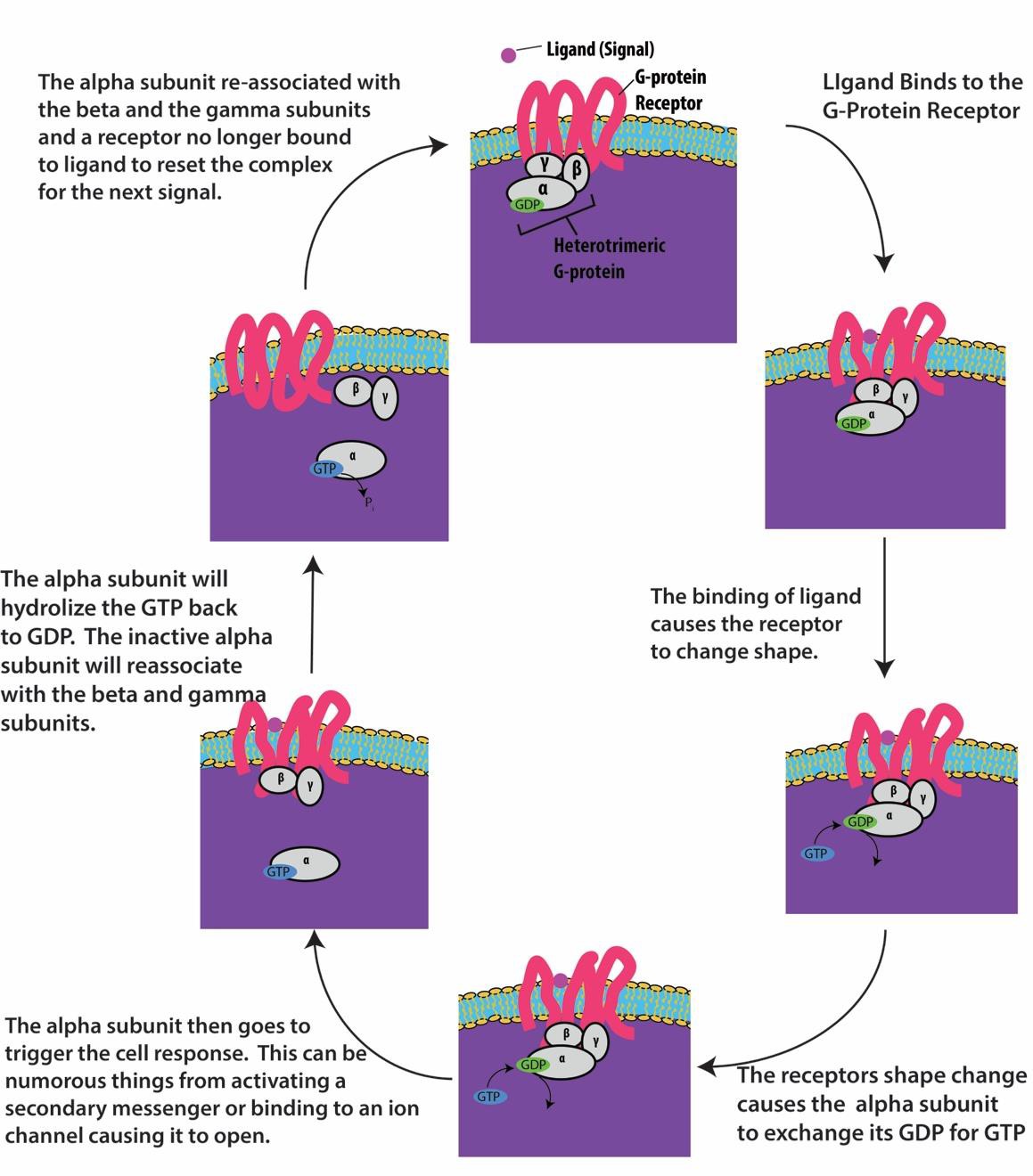
La segnalazione cellulare tramite i recettori legati alle proteine G avviene come un ciclo. Una volta che il ligando si lega al recettore, il cambiamento di forma risultante attiva la proteina G, che rilascia GDP e raccoglie GTP. Le subunità della proteina G si dividono poi in subunità α e βγ. Uno o entrambi questi frammenti di proteina G possono essere in grado di attivare altre proteine nella cellula. Dopo un po’, il GTP sulla subunità α attiva della proteina G viene idrolizzato in GDP e la subunità βγ viene disattivata. Le subunità si riassociano per formare la proteina G inattiva e il ciclo ricomincia (Figura 9.8).
I recettori legati alle proteine G sono usati in molti processi fisiologici, compresi quelli per la trasduzione della visione, il gusto e la regolazione del sistema immunitario e dell’infiammazione.

HER2 è un recettore tirosin-chinasi. Nel 30% dei tumori al seno umani, HER2 è permanentemente attivato, con conseguente divisione cellulare sregolata. Lapatinib, un farmaco usato per trattare il cancro al seno, inibisce il processo con cui il recettore si fosforila, riducendo così la crescita tumorale del 50 per cento. Quale delle seguenti fasi sarebbe inibita da Lapatinib?
- Il legame delle molecole di segnalazione, la dimerizzazione e la risposta cellulare a valle
- Dimerizzazione e la risposta cellulare a valle
- La risposta cellulare a valle
- Attività della fosfatasi, dimerizzazione e la risposta cellulare a valle
9.1.3 Molecole di segnalazione
Prodotti dalle cellule di segnalazione, i ligandi sono segnali chimici che viaggiano verso le cellule bersaglio e causano una risposta. I tipi di molecole che servono come ligandi sono incredibilmente vari e vanno da piccole proteine a piccoli ioni. I ligandi sono classificati come piccoli ligandi idrofobici, che possono attraversare le membrane plasmatiche, o ligandi solubili in acqua, che non possono.
Ligandi idrofobici piccoli
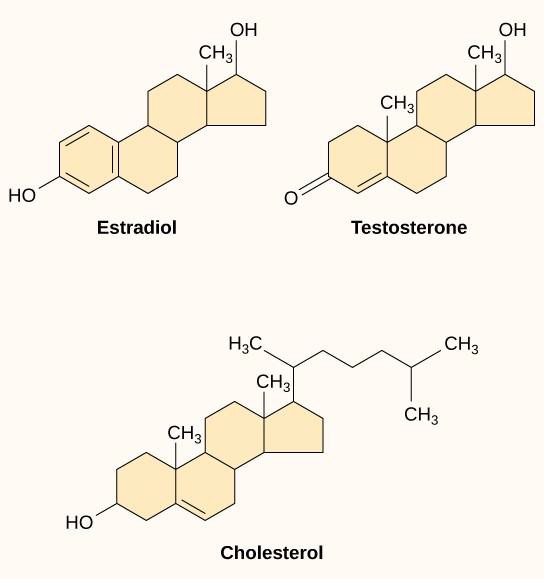
I ligandi idrofobici piccoli, chiamati anche ligandi solubili nei lipidi, possono diffondere direttamente attraverso la membrana plasmatica e interagire con i recettori interni. Membri importanti di questa classe di ligandi sono gli ormoni steroidei. Gli steroidi sono lipidi che hanno uno scheletro idrocarburico con quattro anelli fusi; diversi steroidi hanno diversi gruppi funzionali attaccati allo scheletro di carbonio. Gli ormoni steroidei includono l’ormone sessuale femminile estradiolo, che è un tipo di estrogeno; l’ormone sessuale maschile testosterone; e il colesterolo, che è un importante componente strutturale delle membrane biologiche e un precursore degli ormoni steroidei (Figura 9.10). Altri ormoni idrofobici includono gli ormoni tiroidei e la vitamina D. Per essere solubili nel sangue, i ligandi idrofobici devono legarsi alle proteine di trasporto mentre vengono trasportati attraverso il flusso sanguigno.
Ligandi solubili in acqua
Siccome i ligandi solubili in acqua sono polari, non possono passare attraverso la membrana plasmatica senza aiuto. A volte sono troppo grandi per passare attraverso la membrana. Invece, la maggior parte dei ligandi idrosolubili si lega al dominio extracellulare dei recettori della superficie cellulare (vedi Figura 9.5). Questo gruppo di ligandi è abbastanza vario e include piccole molecole, peptidi e proteine.
9.2 | Propagazione del segnale
Alla fine di questa sezione, sarete in grado di:
- Spiegare come il legame di un ligando inizia la trasduzione del segnale in una cellula.
- Riconoscere che i segnali intracellulari sono trasmessi dal ruolo della fosforilazione o dei secondi messaggeri.
Una volta che un ligando solubile in acqua si lega al suo recettore, il segnale è trasmesso attraverso la membrana e nel citoplasma. La continuazione di un segnale in questo modo è chiamata trasduzione del segnale (Figura 9.5). La trasduzione del segnale avviene solo con i recettori della superficie cellulare, poiché i recettori interni sono in grado di entrare nella cellula.
Quando un ligando si lega al suo recettore, si verificano dei cambiamenti conformazionali che interessano il dominio intracellulare del recettore. Questi cambiamenti conformazionali portano all’attivazione del dominio intracellulare o delle sue proteine associate. In alcuni casi, il legame del ligando causa la dimerizzazione del recettore, il che significa che due recettori si legano tra loro per formare un complesso stabile chiamato dimero. Un dimero è un composto chimico formato quando due molecole (spesso identiche) si uniscono. Il legame dei recettori in questo modo permette ai loro domini intracellulari di entrare in stretto contatto e attivarsi a vicenda.
9.2.1 Vie di segnalazione e amplificazione del segnale
Anche se le molecole di segnalazione si trovano spesso a concentrazioni molto basse, possono produrre effetti profondi. Dopo che il ligando si lega al recettore della superficie cellulare, l’attivazione dei componenti intracellulari del recettore mette in moto una catena di eventi che viene chiamata via di segnalazione o cascata di segnalazione. In una via di segnalazione, secondi messaggeri, enzimi e/o proteine attivate attivano altre proteine o messaggeri (Figura 9.11). Ogni membro della via può attivare migliaia di membri della via successiva in un processo chiamato amplificazione del segnale. Poiché il segnale è amplificato ad ogni passo, una risposta molto grande può essere generata da un singolo recettore che lega un ligando.
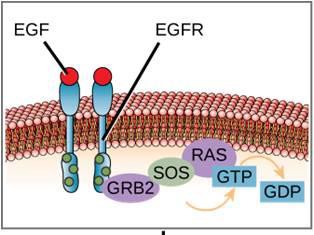
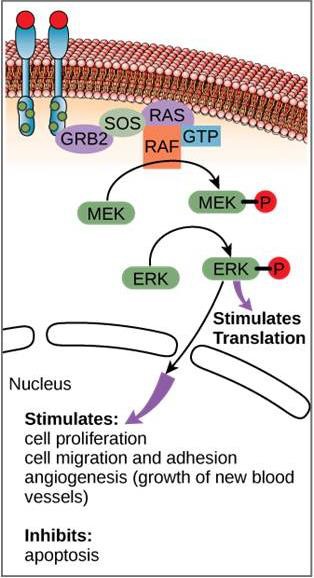
Un esempio di una via di segnalazione è mostrato nella Figura 9.11. Il fattore di crescita epidermico (EGF) è una molecola di segnalazione che è coinvolta nella regolazione della crescita cellulare, nella guarigione delle ferite e nella riparazione dei tessuti. Il recettore per EGF (EGFR) è una tirosina chinasi. Una chinasi attivata fosforila e attiva molte molecole a valle. Quando EGF si lega a EGFR, una cascata di eventi di fosforilazione a valle segnala alla cellula di crescere e dividersi. Se EGFR è attivato in momenti inappropriati, può verificarsi una crescita cellulare incontrollata (cancro).
In certi tipi di cancro, l’attività GTPasi della proteina G RAS è inibita. Questo significa che la proteina RAS non può più idrolizzare il GTP in GDP. Che effetto avrebbe questo sugli eventi cellulari a valle?
9.2.2 Metodi di segnalazione intracellulare
L’induzione di una via di segnalazione dipende dalla modifica di un componente cellulare da parte di un enzima. Ci sono numerose modifiche enzimatiche che possono avvenire per attivare il componente successivo della via. I seguenti sono alcuni degli eventi più comuni nella segnalazione intracellulare.
Fosforilazione
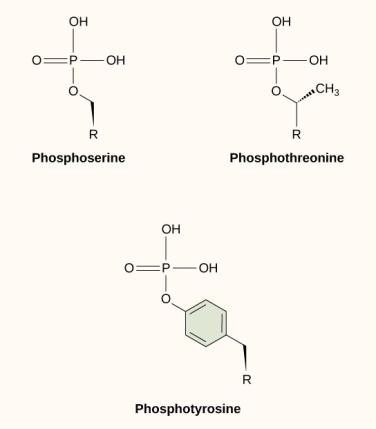
Una delle modifiche chimiche più comuni che avviene nei percorsi di segnalazione è l’aggiunta di un gruppo fosfato a una molecola in un processo chiamato fosforilazione. Il fosfato può essere aggiunto a un nucleotide come il GMP per formare GDP o GTP. I fosfati sono anche spesso aggiunti ai residui di serina, treonina e tirosina delle proteine, dove sostituiscono il gruppo idrossile dell’aminoacido (Figura 9.12). Il trasferimento del fosfato è catalizzato da un enzima chiamato chinasi. La fosforilazione può attivare o inattivare gli enzimi, e l’inversione della fosforilazione, la defosforilazione, invertirà l’effetto.
Secondi messaggeri
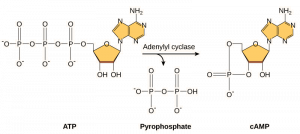
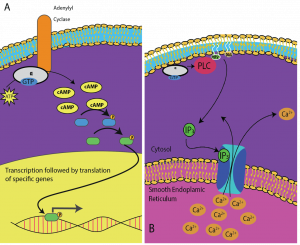
I secondi messaggeri sono piccole molecole che propagano un segnale dopo che è stato iniziato dal legame della molecola di segnalazione al recettore. Queste molecole aiutano a diffondere un segnale attraverso il citoplasma alterando il comportamento di alcune proteine cellulari. Un secondo messaggero utilizzato da diversi tipi di cellule è l’AMP ciclico (cAMP). L’AMP ciclico è sintetizzato dall’enzima adenilciclasi dall’ATP (Figura 9.13). Il ruolo principale del cAMP nelle cellule è quello di legarsi e attivare un enzima chiamato chinasi cAMP-dipendente (A-chinasi). L’A-chinasi regola molte vie metaboliche vitali: Fosforila i residui di serina e treonina delle sue proteine bersaglio, attivandole nel processo. L’A-chinasi si trova in molti tipi diversi di cellule, e le proteine bersaglio in ogni tipo di cellula sono diverse. Un altro messaggero secondario è il Ca2+che può essere rilasciato per inondare la cellula.
Diverse cellule rispondono diversamente al cAMP. Nella Figura 9.14, la subunità alfa di un recettore della proteina G è mostrata mentre attiva due diversi tipi di segnalazione. Nella prima immagine, il cAMP viene prodotto dall’enzima adenilato ciclasi quando viene attivato dalla subunità alfa. Nella seconda immagine, la subunità alfa della proteina G innesca una cascata che rilascia Ca2+ dal reticolo endoplasmatico liscio. In questo caso il Ca2+ è il messaggero secondario che causa la risposta cellulare.
9.3 | Risposta al segnale
Al termine di questa sezione, sarai in grado di:
- Riconoscere che le vie di segnalazione dirigono l’espressione delle proteine, il metabolismo cellulare e la crescita cellulare.
- Riconoscere il ruolo dell’apoptosi nello sviluppo e nel mantenimento di un organismo sano.
Utilizzando le vie di trasduzione del segnale, i recettori nella membrana del plasma producono una varietà di effetti sulla cellula. All’interno della cellula, i ligandi si legano ai loro recettori interni, permettendo loro di influenzare direttamente il DNA della cellula e il meccanismo di produzione delle proteine. I risultati delle vie di segnalazione sono estremamente vari e dipendono dal tipo di cellula coinvolta e dalle condizioni esterne e interne. Un piccolo campione di risposte è descritto di seguito.
9.3.1 Risposte alla via di segnalazione
Espressione genica
Alcune vie di trasduzione del segnale regolano la trascrizione dell’RNA. Altri regolano la traduzione delle proteine.
Aumento del metabolismo cellulare
L’attivazione dei recettori β-adrenergici nelle cellule muscolari da parte dell’adrenalina porta ad un aumento dell’AMP ciclico all’interno della cellula. L’adrenalina è un ormone prodotto dalla ghiandola surrenale che prepara il corpo per emergenze a breve termine.
Crescita cellulare
Le vie di segnalazione delle cellule hanno anche un ruolo importante nella divisione cellulare. Le cellule normalmente non si dividono se non sono stimolate da segnali provenienti da altre cellule. I ligandi che promuovono la crescita cellulare sono chiamati fattori di crescita. La maggior parte dei fattori di crescita si lega ai recettori della superficie cellulare che sono collegati alle tirosin-chinasi.
Morte cellulare
Quando una cellula è danneggiata, superflua, o potenzialmente pericolosa per un organismo, una cellula può avviare un meccanismo per innescare la morte cellulare programmata, o apoptosi. L’apoptosi permette ad una cellula di morire in un modo controllato che impedisce il rilascio di molecole potenzialmente dannose dall’interno della cellula. Tuttavia, in alcuni casi, come un’infezione virale o una divisione cellulare incontrollata dovuta al cancro, i normali controlli ed equilibri della cellula falliscono. Anche la segnalazione esterna può avviare l’apoptosi. Per esempio, la maggior parte delle cellule animali normali hanno recettori che interagiscono con la matrice extracellulare, una rete di glicoproteine che fornisce supporto strutturale alle cellule animali. Il legame dei recettori cellulari alla matrice extracellulare avvia una cascata di segnalazione all’interno della cellula. Tuttavia, se la cellula si allontana dalla matrice extracellulare, la segnalazione cessa e la cellula va incontro ad apoptosi. Questo sistema aiuta a prevenire che le cellule viaggino attraverso il corpo e proliferino fuori controllo, come succede con le cellule tumorali che metastatizzano.
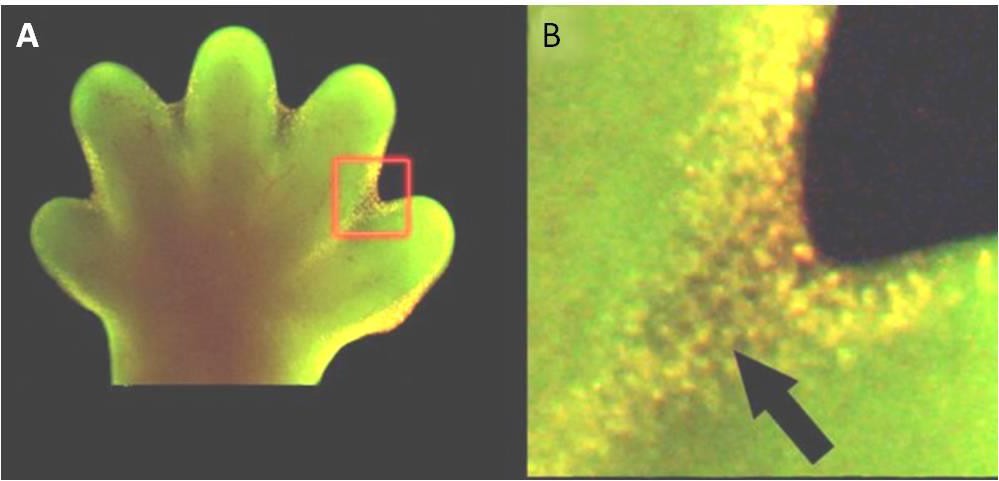
L’apoptosi è anche essenziale per il normale sviluppo embriologico. Nei vertebrati, per esempio, le prime fasi dello sviluppo includono la formazione di tessuto simile a una ragnatela tra le singole dita delle mani e dei piedi (Figura 9.15). Nel corso del normale sviluppo, queste cellule non necessarie devono essere eliminate, permettendo la formazione di dita delle mani e dei piedi completamente separate. Un meccanismo di segnalazione cellulare innesca l’apoptosi, che distrugge le cellule tra le dita in via di sviluppo.
9.3.2 Cessazione delle vie di segnalazione
La segnalazione aberrante spesso osservata nelle cellule tumorali è la prova che la cessazione di un segnale al momento opportuno può essere importante quanto l’inizio di un segnale. Un metodo per fermare un segnale specifico è quello di degradare il ligando o rimuoverlo in modo che non possa più accedere al suo recettore. Uno dei motivi per cui gli ormoni idrofobici come gli estrogeni e il testosterone innescano eventi di lunga durata è perché si legano a proteine carrier. Queste proteine permettono alle molecole insolubili di essere solubili nel sangue, ma proteggono anche gli ormoni dalla degradazione da parte degli enzimi circolanti.
All’interno della cellula, molti enzimi diversi invertono le modifiche cellulari che risultano dalle cascate di segnalazione. Per esempio, le fosfatasi sono enzimi che rimuovono il gruppo fosfato attaccato alle proteine dalle chinasi in un processo chiamato defosforilazione. Il cAMP viene degradato in AMP dalla fosfodiesterasi, terminando così il suo segnale. Allo stesso modo, il rilascio delle riserve di calcio è invertito dalle pompe di Ca2+ che si trovano nelle membrane cellulari.