Détail de l’expérience
- Introduction
- Matériels et méthodes
- Questions de réflexion et de discussion
- Références et liens
- Outils d’évaluation des résultats d’apprentissage des étudiants
- . de réflexion et de discussion
- Références et liens
- Outils d’évaluation des résultats d’apprentissage des étudiants
- Outils d’évaluation formative de cette expérience
Introduction :
Dans cette enquête, nous examinerons les effets de la compétition pour les ressources sur la production reproductive au sein et entre deux espèces de guêpes parasitoïdes.
Objectifs du laboratoire :
A la fin de ce laboratoire, les élèves seront en mesure de….
- Décrire le cycle de vie de Nasonia vitripennis et Melittobia digitata,
- Expliquer les interactions possibles entre deux espèces de parasites en compétition pour la même ressource hôte,
- Concevoir une expérience pour déterminer la nature de l’interaction entre ces deux espèces lorsqu’elles sont en compétition pour un hôte commun,
- Mener une expérience consensuelle pour déterminer les effets de la compétition intraspécifique et interspécifique sur la production reproductive chez Nasonia vitripennis et Melittobia digitata,
- Utiliser les données résultantes pour estimer les paramètres du modèle de compétition Lotka-Volterra,
- Relier les résultats de la recherche en classe au principe d’exclusion de la compétition.
Les communautés écologiques sont composées de populations de toutes les espèces dans un habitat. La structure d’une communauté sera déterminée en partie par la dynamique des interactions entre les espèces de la communauté. Les interactions entre deux espèces peuvent être directes ou indirectes (c’est-à-dire médiées par d’autres espèces).
Dans une communauté naturelle même simple, des centaines d’espèces différentes de plantes et d’animaux interagissent les unes avec les autres. Cependant, malgré cette diversité, nous pouvons identifier des catégories d’interactions qui ont des effets différents sur la croissance de la population (tableau 1). Ces catégories sont définies par la direction des effets sur les espèces en interaction.
En plus des interactions entre les espèces, les interactions entre les individus d’une même espèce peuvent également être importantes pour structurer une communauté. Les interactions intra-spécifiques peuvent affecter la dynamique de la population de l’espèce, qui à son tour influencera les interactions entre les espèces. La compétition intraspécifique se produit lorsque différents individus d’une même espèce ou d’une même population se disputent une ressource. Ces interactions peuvent être féroces car les individus ont besoin des mêmes ressources limitées pour survivre et se reproduire. Lorsque différentes espèces se disputent la même nourriture, le même habitat ou une autre ressource environnementale, on parle de compétition interspécifique. Ces interactions sont généralement un peu moins intenses. En effet, si les besoins de deux espèces peuvent être similaires, ils ne peuvent jamais être aussi proches que pour les individus d’une même espèce.
| Tableau 1. Catégories d’interactions directes entre deux espèces dans la même communauté | |||||
| Nom de l’interaction | Type de contact | Effet direct sur l’espèce #1 | Effet direct sur l’espèce #2 | . effet sur l’espèce n°2 | Autres aspects de la relation |
| Relation neutre | Deux espèces ne sont liées qu’indirectement par des interactions avec d’autres espèces. | 0 | 0 | Chaque espèce a une relation neutre avec la plupart des espèces de son habitat. | |
| Commensalisme | Relation qui aide directement une espèce mais qui n’affecte pas beaucoup l’autre, voire pas du tout. | + | 0 | Le commensalisme, le mutualisme et le parasitisme sont tous des cas de symbiose. | |
| Mutualisme | Les bénéfices circulent dans les deux sens entre les espèces en interaction. | + | Mieux vu comme une exploitation à double sens que comme une coopération douillette. | ||
| Prédation Vrais prédateurs Brouteurs |
Le prédateur attaque et se nourrit d’une série de proies mais ne s’installe pas dans ou sur elles. | + | La proie meurt généralement. Avec les brouteurs, la plante peut ou non mourir. | ||
| Parasitisme Parasites Parasitoïdes | Le parasite se nourrit des tissus d’un ou plusieurs hôtes, résidant dans ou sur eux pendant au moins une partie de son cycle de vie. | + | Un hôte peut ou non mourir à la suite de l’interaction. | ||
| Concurrence interspécifique | Les inconvénients peuvent circuler dans les deux sens entre les espèces, ou le concurrent supérieur peut être largement épargné. | Généralement moins intense que la concurrence entre membres de la même espèce. | |||
0 signifie aucun effet direct sur la croissance de la population.
+ signifie un effet positif ; signifie un effet négatif.
Considérez, cependant, le cas théorique de deux espèces qui occupent la même niche. Gause (1934) a étudié deux espèces de protistes qui se nourrissaient toutes deux des mêmes cellules bactériennes. Lorsqu’il les a combinées dans une seule culture, l’une a toujours poussé l’autre à l’extinction. Depuis, de nombreuses autres expériences ont confirmé la « loi de Gause », aujourd’hui appelée « principe d’exclusion compétitive ». Elle stipule que deux espèces qui utilisent des ressources identiques ne peuvent coexister indéfiniment ou que « des concurrents complets ne peuvent coexister » (Hardin 1960).
De nombreuses expériences ont démontré que plus deux espèces dans un habitat diffèrent dans leur utilisation des ressources, plus il est probable qu’elles puissent, en fait, coexister (Krebs 1994). Même deux espèces qui se chevauchent beaucoup peuvent vivre ensemble pendant un certain temps, bien que les interactions compétitives suppriment souvent le taux de croissance de l’une ou des deux espèces. Au fil du temps, un phénomène intéressant appelé partitionnement des ressources peut se produire. Les membres de chaque espèce peuvent en venir à se spécialiser dans une subdivision d’une catégorie de ressources similaires. Par exemple, si les deux se nourrissent de pommes, l’un peut se nourrir de petits fruits verts et l’autre de fruits plus gros et plus mûrs.
Le modèle Lotka-Volterra a été développé pour permettre aux écologistes de prédire le résultat potentiel lorsque deux espèces sont en compétition pour les mêmes ressources. Fondamentalement, le modèle tente de rendre compte de l’effet que la présence d’une espèce aura sur la croissance de la population de l’autre espèce, par rapport à l’effet de compétition que deux membres de la même espèce auraient l’un sur l’autre.
L’équation pour la croissance de la population de l’espèce 1 est:
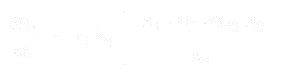
Et pour l’espèce 2, elle est:
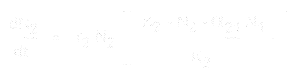
où :
- * N1 et N2 sont les tailles de population des espèces 1 et 2,
* r1 et r2 sont les taux d’accroissement intrinsèques de ces espèces,
* K1 et K2 sont les capacités de charge de l’habitat pour chaque espèce,
* alpha12 et alpha21 sont les effets d’une espèce sur la croissance de la population de l’autre. Plus précisément, alpha12 est l’effet de l’espèce 2 sur la croissance de l’espèce 1, et alpha21 est l’effet de l’espèce 1 sur la croissance de l’espèce 2.
Si les valeurs de chaque équation sont connues (ou peuvent être estimées empiriquement à partir des résultats d’une expérience), alors l’équation peut être utilisée pour prédire le résultat potentiel d’une compétition (c’est-à-dire si elles peuvent coexister ou si l’une finira par exclure l’autre). Les valeurs de K1, K2, a12 et a21 sont utilisées pour tracer les isoclines de croissance nulle (c’est-à-dire lorsque dN1/dt ou dN2/dt est égal à zéro) pour les deux espèces sur le même graphique, et les sommes résultantes des vecteurs de croissance de la population (trajectoires) sont utilisées pour déterminer le résultat de la compétition (Figure 1).

Figure 1. Exemples de graphiques d’isoclines de croissance nulle pour lesquelles les espèces 1 et 2 coexistent (à gauche), et l’espèce 1 exclut compétitivement l’espèce 2 (à droite).
Le modèle de compétition Lotka-Volterra décrit le résultat de la compétition entre deux espèces au cours du temps écologique. Parce qu’une espèce peut exclure de manière compétitive une autre espèce (figure 1) dans le temps écologique, l’espèce inférieure sur le plan compétitif peut augmenter la gamme des types de nourriture qu’elle consomme pour survivre. Cependant, la réponse des espèces à la compétition interspécifique au cours de l’évolution est souvent l’inverse de ce qui se passe en temps écologique. Les compétiteurs se spécialisent généralement sur des types de ressources particuliers. Cette répartition des ressources qui se produit au cours de l’évolution entraîne en fait une diminution ou l’absence de compétition entre les deux espèces.
Bien qu’elles ne soient pas particulièrement proches les unes des autres, les histoires de vie de deux espèces de guêpes parasitoïdes, Melittobia digitata et Nasonia vitripennis, sont assez similaires. Les deux espèces sont capables d’utiliser le même hôte, bien que dans la nature elles utilisent des hôtes différents. Les Melittobia sont environ deux fois moins grandes que les Nasonia, mais toutes deux sont assez petites et totalement inoffensives pour les humains.
Leurs cycles de vie complets sont relativement courts (2-4 semaines à 25o C), et également assez similaires (Figure 2). Les femelles pondent de nombreux œufs à travers le revêtement de l’hôte. Les œufs éclosent pour devenir des larves qui consomment l’hôte, puis se transforment en nymphes, et enfin se métamorphosent en un stade adulte. Chez Melitobbia digitata, les femelles adultes peuvent avoir des ailes normales ou rabougries. Les adultes à ailes normales se dispersent à partir de l’hôte pour chercher de nouvelles ressources alimentaires. Les femelles sans ailes pondront leurs œufs sur le même hôte dont elles ont émergé, ou se disperseront vers un nouvel hôte dans le même nid (Freeman et Ittyeipe 1976, Côsoli et Vinson 2002).
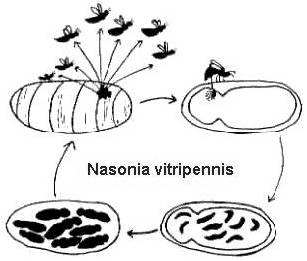
Figure 2. Le cycle de vie de Nasonia vitripennis sur une chrysalide hôte de Neobellieria bullata (dessin de Bethia King). Le cycle de vie de Melittobia digitata est le même, bien que les individus à tous les stades soient plus petits.
Il existe environ 70 000 espèces connues de parasitoïdes dans le monde (9% de tous les insectes), mais les estimations de leur nombre vont jusqu’à 800 000 (Strand 2002). Parmi les espèces décrites, environ 80 % sont des membres de l’ordre des hyménoptères, comme les deux espèces que vous utilisez dans ce laboratoire. Dans la nature, les hôtes peuvent être parasités par plus d’une espèce de parasitoïde, toutes en compétition pour la même ressource (Stand 2002). Comme l’espèce hôte peut elle-même être un parasite d’une autre espèce, les effets écologiques de cette compétition sur la structure de la communauté peuvent être très complexes. Par exemple, le diprion du pin gris (Neodiprion swainei Midd.), qui peut attaquer, défolier et tuer de grands peuplements de pins gris en Amérique du Nord, est l’hôte de 11 espèces différentes de guêpes parasitoïdes. Il semble que la plupart de ces espèces coexistent parce qu’elles se partagent la ressource hôte en parasitant différents stades du cycle de vie de l’hôte, ou différents segments de la population hôte (Price 1972). Un membre de la guilde des parasitoïdes qui s’attaque aux cocons des tenthrèdes est une espèce introduite (Pleolophus basizonus). C’est un compétiteur supérieur et sa présence détermine l’abondance de certaines de ses espèces concurrentes (Price 1970). Bien que la compétition puisse affecter la structure des communautés de parasitoïdes, la variation spatiale et temporelle des ressources de l’hôte peut être encore plus importante pour déterminer la richesse des espèces de parasitoïdes, même en présence d’une compétition interspécifique entre parasitoïdes (Hawkins 2000).
Les parasitoïdes dont les hôtes sont des ravageurs importants des cultures ou des arbres forestiers sont parfois lâchés intentionnellement comme contrôles biologiques des populations de ravageurs. Lorsque plus d’une espèce de parasitoïde s’attaque au même hôte, il est important de comprendre la nature de cette compétition avant de procéder à des lâchers. Si les différentes espèces de parasitoïdes sont capables de coexister grâce à la répartition des ressources, le contrôle de l’hôte nuisible peut être mieux réalisé en introduisant certaines ou toutes les espèces de parasitoïdes. D’autre part, si les concurrents limitent leurs populations respectives en raison de leurs interactions compétitives, alors le contrôle maximal de l’espèce nuisible pourrait être atteint en libérant la plus efficace des espèces de parasitoïdes par elle-même (Amarasekare 2000).
Matériels et méthodes :
Aperçu des méthodes de collecte et d’analyse des données :
Lab 1 :
Les catégories d’interactions abordées dans le tableau 1 peuvent sembler assez simples lorsqu’on ne fait que les lire. Mais si vous deviez observer deux animaux non familiers en train d’interagir, comment décideriez-vous de l' »étiquette » à appliquer ? Pourriez-vous prédire le résultat de l’interaction ? Comment pourriez-vous tester votre prédiction ?
Les deux guêpes parasitoïdes présentées dans cette enquête de laboratoire semblent occuper des niches similaires. Nous sommes intéressés par les interactions entre les deux espèces. De plus, nous voulons être en mesure de quantifier l’effet d’une espèce sur l’autre.
Chaque groupe doit :
- – discuter et énumérer toutes les combinaisons expérimentales possibles qui pourraient être mises en place impliquant deux guêpes parasites, Melittobia digitata et Nasonia vitripennis, et un seul hôte, Neobellierria,
– prédire ce que vous pensez être le résultat pour chaque interaction possible,
– identifiez et listez les variables que vous manipuleriez dans votre expérience,
– identifiez et listez les variables que vous garderiez constantes dans votre expérience,
– identifiez et listez les variables dépendantes que vous voudriez mesurer pour quantifier l’effet de chaque espèce sur l’autre.
Chaque groupe partagera son plan expérimental avec le reste de la classe. Ensemble, en tant que classe, nous développerons un plan expérimental consensuel. Sur la base du plan expérimental consensuel, chaque personne doit mettre en place une culture répétée pour chaque traitement. Voir « Manipulation des parasitoïdes » ci-dessous. Nous mettrons en commun les données de toute la classe pour les analyser.
Contrôles hebdomadaires :
Chaque semaine, chaque personne devrait inspecter ses cultures pour voir si des guêpes adultes ont émergé. Vous devriez enregistrer la date à laquelle vous voyez pour la première fois des adultes émergés pour chaque culture. Les cultures de Nasonia doivent être congelées 21 jours après avoir été établies. Les cultures de Melittobia et d’espèces mixtes doivent être congelées après 42 jours.
Laboratoire 2 (6 semaines après le laboratoire 1) :
Chaque personne doit compter le nombre de descendants produits dans chaque culture répliquée. Entrez vos données dans une feuille de calcul afin que les données de la classe puissent être regroupées. Utilisez les données regroupées pour estimer les paramètres du modèle de compétition Lotka-Volterra et pour l’analyse statistique des effets de la compétition.
Dans vos groupes, discutez de la façon dont les données peuvent être utilisées pour quantifier les paramètres du modèle de compétition de Lotka-Volterra (voir « Quantifier le modèle de compétition de Lokta-Volterra » ci-dessous). Discutez également de ce que les comparaisons de traitements particuliers peuvent nous apprendre sur l’importance relative de la compétition intraspécifique et interspécifique chez ces deux espèces de parasitoïdes (voir « Directives pour l’analyse des données » ci-dessous).
Manipulation des parasitoïdes :
Les adultes des deux parasitoïdes, Melittobia digitata et Nasonia vitripennis, sont très « conviviaux ». Bien que les femelles possèdent des ailes normales et puissent voler, elles ne le font pas facilement. Cependant, elles sont négativement géotaxiques (c’est-à-dire qu’elles se déplacent vers le haut, loin de la gravité). Lorsque quelques femelles d’une culture sont secouées sur une surface horizontale, puis recouvertes d’un flacon de verre inversé, elles se hissent facilement dans le flacon et sur les côtés. Une fois que vous avez des guêpes dans une fiole, vous pouvez facilement ajouter une chrysalide hôte, puis boucher la fiole hermétiquement avec du coton. Cette méthode permet de manipuler efficacement un grand nombre d’individus. Les guêpes adultes peuvent également être manipulées avec des cure-pipes courts, auxquels les guêpes vont temporairement adhérer.
Quantification du modèle de compétition Lotka-Volterra :
Le modèle de compétition de Lotka-Volterra a été décrit et défini avec des équations dans l’introduction. Comme indiqué là, si les valeurs de chaque équation peuvent être estimées empiriquement à partir des résultats d’une expérience, alors l’équation peut être utilisée pour prédire le résultat potentiel d’une compétition (c’est-à-dire si les deux espèces coexisteront ou si l’une finira par exclure l’autre). Les valeurs de K1, K2, alpha12 et alpha21 sont utilisées pour tracer les isoclines de croissance nulle (c’est-à-dire lorsque dN1/dt ou dN2/dt est égal à zéro) pour les deux espèces sur le même graphique, et les sommes résultantes des vecteurs de croissance de la population (trajectoires) sont utilisées pour déterminer le résultat de la compétition.
Sur la base de notre plan expérimental, nous devons déterminer les valeurs de ces paramètres. Rappelons que la capacité de charge d’une population est le nombre maximum d’individus qui peuvent survivre dans un habitat. Pour simplifier cette expérience, nous avons défini l’habitat des parasitoïdes comme étant un seul hôte. Dans la réalité, bien sûr, un habitat contiendrait probablement plus d’un nid de larves ou d’un puparium de mouche à viande, et il y aurait donc de nombreux hôtes potentiellement exploitables. Pour déterminer les capacités de charge des deux espèces, nous devons connaître le nombre maximum de descendants d’une espèce donnée qui peuvent être produits sur un seul hôte lorsque seule cette espèce est présente. En gardant cela à l’esprit, les données de quel traitement seraient utilisées pour estimer les capacités de charge de Melittobia et Nasonia ? (Rappelez-vous qu’à la capacité de charge, toutes les ressources de l’hôte seront utilisées.)
L’estimation des coefficients de compétition (alpha12, et alpha21) est un peu plus compliquée. Rappelons que l’équation pour la croissance de la population de l’espèce 1 est :
Lorsque toute la ressource de l’hôte est utilisée par les parasitoïdes, alors une population ne peut plus croître. En d’autres termes, dN1 /dt = 0. Cette condition se produira lorsque K1 – N1 – alpha12*N2 = 0. Pour trouver alpha12, nous devons le résoudre (c’est-à-dire faire un peu d’algèbre) et ensuite substituer des valeurs pour K1, N1 et N2. Nous avons décrit plus haut comment trouver les capacités de charge. En supposant que le Melittobia est « l’espèce 1 », utilisez sa capacité de charge pour K1. Le nombre de descendants de Melittobia et de Nasonia produits dans la compétition interspécifique est N1 et N2, respectivement. Dans cette optique, les données de quel traitement seraient utilisées pour estimer N1 et N2 ?
La même approche que vous avez utilisée pour calculer alpha12, peut être utilisée pour calculer alpha21.
Maintenant que vous avez calculé toutes les valeurs des paramètres, vous pouvez utiliser ces valeurs pour tracer les isoclines de croissance nulle et prédire le résultat de la compétition entre Melittobia et Nasonia.
Lignes directrices pour l’analyse des données :
Nous pouvons utiliser les comparaisons entre différents traitements pour explorer l’importance relative de la compétition intraspécifique et interspécifique. Tout d’abord, identifiez quel type de compétition, intraspécifique ou interspécifique, le cas échéant, se produit dans chaque traitement. Après avoir fait cela, réfléchissez à toutes les comparaisons entre les paires de traitements. Que nous apprend chacune de ces comparaisons ? Il pourrait être utile de produire un tableau qui énumère les comparaisons et leur signification. Puisque toutes les comparaisons impliquent deux traitements, elles peuvent être analysées statistiquement à l’aide de tests t.
Questions pour approfondir la réflexion et la discussion:
- Sur la base des valeurs des paramètres que vous avez calculées pour le modèle de compétition Lotka-Volterra, quel est le résultat prédit de la compétition entre les deux espèces ? Le résultat prédit a-t-il été atteint dans chaque réplique de la compétition interspécifique ? Si non, pourquoi ?
- La « loi de Gause » stipule que les concurrents qui partagent exactement les mêmes ressources de la même manière ne peuvent pas coexister. Cela signifie que l’espèce qui utilise le plus efficacement la ressource contestée finira par éliminer l’autre à cet endroit. La loi de Gause semble-t-elle s’appliquer à l’interaction entre Melittobia et Nasonia ? Pourquoi ou pourquoi pas ?
- Si ces deux espèces utilisaient le même hôte dans la nature, comment le partage des ressources pourrait-il leur permettre de coexister ?
- Selon les résultats de votre expérience, pourquoi les deux espèces n’utilisent-elles pas le même hôte dans la nature ?
- Selon les valeurs estimées des capacités de charge et des coefficients de compétition, prédisez le résultat de la compétition entre Melittobia et Nasonia en utilisant le modèle de compétition Lotka-Volterra chez Populus (voir Références et liens). Le résultat prédit de la compétition est-il affecté par la taille initiale de la population ou le taux de croissance de la population ? Si oui, comment ? Comment le temps pour atteindre l’équilibre est-il affecté par ces valeurs ?
- Les capacités de charge et les coefficients de compétition ne sont que des estimations. Quels facteurs pourraient affecter les capacités de charge et les coefficients de compétition pour ces deux espèces ?
- Si une compétition interspécifique se produit chez ces espèces, comment pourrions-nous déterminer quel mécanisme de compétition (interférence ou exploitation) se produit ?
*** Note : Les réponses à plusieurs de ces questions et de nombreux autres commentaires de l’auteur contributeur se trouvent dans les « NOTES AU FACULTÉ : Commentaires sur les questions pour une réflexion plus approfondie ».
Références et liens:
- Abe, J., Y. Kamimura, N. Kondo, et M. Shimada. 2003. Ratio sexuel extrêmement biaisé par les femelles et combat létal entre mâles chez une guêpe parasitoïde, Melittobia australica (Eulophidae). Behavioural Ecology 14 : 34-39.
- Amarasekare, P. 2000. Coexistence de parasitoïdes concurrents sur un hôte distribué de façon inégale : mécanismes locaux vs spatiaux. Ecologie 81 : 1286-96.
- Cooperband M. F., R. W. Matthews, S. B. Vinson. 2003. Les facteurs affectant la biologie de la reproduction de Melittobia digitata (Hymenoptera : Eulophidae) et l’échec à répondre aux prédictions de sex-ratio de la théorie de la concurrence locale des compagnons de Hamilton. Entomologia Experimentalis et Applicata 109 : 1-12.
- Côsoli, F. L., et S. B. Vinson. 2002. Taille de la ponte, développement et différenciation de la morphologie des ailes de Melittobia digitata. Entomologia Experimentalis et Applicata 102 : 135-143.
- Freeman, B. E., et K. Ittyeipe. 1976. Études de terrain sur la réponse cumulative de Melittobia sp. complexe Hawaiiensis) (Eulophidae) à des densités d’hôtes variables. Journal of Animal Ecology 45 : 415-423.
- Gause, G. F. 1934. La lutte pour l’existence. Williams et Wilkins, Baltimore.
- Harden, G. 1960. Le principe d’exclusion compétitive. Science 131 : 1292-1297.
- Hawkins, B. A. 2000. La coexistence des espèces dans les communautés de parasitoïdes : la concurrence importe-t-elle ? Pages 198-213 dans M. E. Hochberg et A. R. Ives (eds). Parasitoid Population Biology. Princeton University Press, Princeton, New Jersey.
- King, B. 2000. Nasonia vitripennis, la guêpe bijoutière. Illustrer les concepts de science et de biologie avec un insecte. www.bios.niu.edu/bking/nasonia.htm
- Krebs, C. J. 1994. Ecologie. L’analyse expérimentale de la distribution et de l’abondance. 4e édition. HarperCollins College Publishers, New York, NY.
- Mackauer, M., B. Bai, A. Chow, et T. Danyk. 1992. La concurrence larvaire asymétrique entre deux espèces de guêpes parasitoïdes solitaires : l’influence de superparasititsm. Entomologie écologique 17 : 233-236.
- Price, P. 1970. Parasitoïdes utilisant le même hôte : nature adaptative des différences de taille et de forme. Ecology 53 : 445-454.
- Price, P. 1972. Caractéristiques permettant la coexistence entre les parasitoïdes d’une tenthrède au Québec. Écologie 51 : 190-195.
- Strand, M. R. 2002. Les interactions entre les parasitoïdes au stade larvaire et leurs hôtes. Pages 129-152 dans E. E. Lewis, J. F. Campbell, et M. V. K. Sukhdeo (eds). L’écologie comportementale des parasites. CABI Publishing, New York.
- Silva-Torres, C., et R. W. Matthews. 2003. Développement de Melittobia australica Girault et M. digitata Dahms (Hymenoptera : Eulophidae) parasitant les puparia de Neobellieria bullata (Parker) (Diptera : Sarcophagidae). Neotropical Entomology 32 : 645-651.
- Werren, J. H. 1983. L’évolution du sex-ratio sous la concurrence locale des compagnons chez une guêpe parasite. Evolution 37:116-124.
- Werren, J.H. 1984. Taille des couvées et régulation du sex-ratio chez la guêpe parasite Nasonia vitripennis (Walker) (Hymenoptera : Pteromalidae). Journal néerlandais de zoologie 34 : 123-143.
Pour des informations sur l’histoire naturelle et les habitats, visitez :
- www.wowbugs.com pour Melittobia,
www.bios.niu.edu/bking/nasonia.htm pour Nasonia,
www.rochester.edu/College/BIO/labs/WerrenLab/nasonia/ pour Nasonia.
Populus peut être téléchargé sur www.cbs.umn.edu/populus.
Outils d’évaluation des résultats d’apprentissage des étudiants :
L’évaluation a été réalisée de diverses manières dans les différents établissements qui ont utilisé cet exercice. Dans tous les cas, les étudiants sont évalués sur la base d’un article scientifique rédigé par chaque étudiant individuellement, ou par les étudiants en groupe. Dans certains cas, les étudiants sont évalués sur la première et la deuxième version d’un article.
La rubrique de notation des articles varie selon l’instructeur. Vous trouverez ci-dessous un exemple de rubrique d’évaluation utilisée au Morehouse College pour un » résumé des résultats « , qui comporte toutes les composantes d’un article scientifique, à l’exception des méthodes. Dans cette rubrique d’évaluation, le « public » concerne le choix du public approprié par l’étudiant. Les étudiants doivent rédiger leur rapport comme s’il s’agissait d’un article scientifique. Le public approprié est donc celui des pairs qui n’ont pas réalisé l’expérience mais qui ont des connaissances scientifiques. Les rapports écrits à l’intention de l’instructeur ou des autres élèves de la classe ne constituent pas un public approprié. Le « format » est l’organisation générale du rapport en sections qui ont une organisation parallèle et s’appuient les unes sur les autres. Par exemple, la discussion doit évaluer les résultats présentés dans la section Résultats et les placer dans un contexte plus large. La Discussion doit également aborder l’hypothèse énoncée dans l’Introduction.
| Évaluation sommaire des résultats (50 points possibles) | |
| Introduction et page de titre (10 points) | _____ |
| Résultats (10 points) | _____ |
| Discussion et conclusions (10 points) | _____ |
| Utilisation de la littérature et citations (10 points) | _____ |
| Format, Audience (10 points) | _____ |
Au niveau de l’introduction, les étudiants remettent tous les tableaux générés par la classe, les réponses aux questions de discussion et les notes hebdomadaires sur l’avancement de l’enquête. En plus des articles scientifiques individuels, les étudiants pourraient présenter les résultats de l’expérience sous la forme d’articles scientifiques de groupe, de présentations orales de groupe ou de présentations d’affiches de groupe. Cependant, puisque tous les élèves réalisent la même expérience et présentent donc les mêmes résultats, les articles scientifiques individuels ou de groupe seraient les plus efficaces.
Outils d’évaluation formative de cette expérience :
Dans le cours d’écologie du Morehouse College, on demande à chaque étudiant d’évaluer chaque exercice de laboratoire sur une échelle de 10 points en fonction de l’efficacité perçue de chaque étude pour renforcer leurs connaissances et leur compréhension des sujets abordés dans le cours magistral d’écologie. En outre, on demande aux étudiants quels exercices ont été les moins et les plus agréables et lesquels ont le plus renforcé leur compréhension de la méthode scientifique. Ces informations sont utilisées pour choisir et modifier les exercices utilisés lors des semestres suivants.
Dans les cours d’écologie de l’Université Emory et de l’Université Radford, on demande à chaque étudiant quels sont les exercices qu’il a préférés et ceux qu’il a le moins appréciés. Les étudiants fournissent parfois des commentaires sur les raisons pour lesquelles ils ont évalué les exercices de la manière dont ils l’ont fait. Là encore, ces informations sont utilisées pour modifier les exercices utilisés lors des semestres suivants.
Une discussion approfondie sur l’évaluation figure dans la section Enseignement de ce site.