
Typ dina exempel här.
- 9.1 Signalmolekyler och cellulära receptorer
- 9.2 Signalens spridning
- 9.3 Reaktion på signalen
Föreställ dig hur livet skulle se ut om du och människorna i din omgivning inte kunde kommunicera. Du skulle inte kunna uttrycka dina önskemål till andra och du skulle inte heller kunna ställa frågor för att få reda på mer om din omgivning. Den sociala organisationen är beroende av kommunikation mellan de individer som ingår i det samhället.
Som för människor är det viktigt för enskilda celler att kunna interagera med sin omgivning och med varandra. Detta gäller oavsett om en cell växer av sig själv i en damm eller om den är en av många celler som bildar en större organism. För att kunna reagera korrekt på externa stimuli har cellerna utvecklat komplexa kommunikationsmekanismer så att de kan ta emot ett meddelande, överföra informationen över plasmamembranet och sedan producera förändringar inom cellen som svar på meddelandet.
I flercelliga organismer sänder och tar cellerna ständigt emot kemiska meddelanden för att samordna andra organs, vävnaders och cellers åtgärder. Förmågan att skicka meddelanden snabbt och effektivt gör det möjligt för cellerna att samordna och finjustera sina funktioner. Även om behovet av cellulär kommunikation i större organismer verkar uppenbart, kommunicerar även encelliga organismer med varandra. Jästceller signalerar till varandra för att underlätta parningen. Vissa former av bakterier samordnar sina handlingar för att bilda stora komplex som kallas biofilmer eller för att organisera produktionen av gifter för att avlägsna konkurrerande organismer. Cellernas förmåga att kommunicera genom kemiska signaler har sitt ursprung i enskilda celler och var avgörande för utvecklingen av flercelliga organismer. Effektiv, felfri kommunikation är avgörande för allt liv.
- 9.1 | Signalmolekyler och cellulära receptorer
- 9.1.1 Former av signalering
- Paracrin signalering
- Autokrin signalering
- Endokrin signalering
- Direkt signalering
- 9.1.2 Typer av receptorer
- Interna receptorer
- Cellytoreceptorer
- 9.1.3 Signalmolekyler
- Små hydrofoba ligander
- Vattenlösliga ligander
- 9.2 | Signalens utbredning
- 9.2.1 Signalvägar och signalförstärkning
- 9.2.2 Metoder för intracellulär signalering
- Fosforylering
- Sekundbudbärare
- 9.3 | Svar på signalen
- 9.3.1 Reaktioner på signalvägen
- Genuttryck
- Ökning av cellmetabolismen
- Celltillväxt
- Celldöd
- 9.3.2 Avslutande av signalvägar
9.1 | Signalmolekyler och cellulära receptorer
I slutet av det här avsnittet kommer du att kunna:
- Beskriv fyra typer av signalering som finns i flercelliga organismer.
- Genomföra interna receptorer med receptorer på cellytan.
- Känn igen sambandet mellan en ligands kemi och dess verkningsmekanism.
Det finns två typer av kommunikation i de levande cellernas värld. Kommunikation mellan celler kallas intercellulär signalering och kommunikation inom en cell kallas intracellulär signalering. Ett enkelt sätt att komma ihåg skillnaden är att förstå att prefixet inter- betyder ”mellan” (en interstatlig motorväg går mellan olika delstater) och intra- betyder ”inuti” (ett dropp betyder intravenöst eller ”inuti venen”).
Kemiska signaler avges av en signalcell och tas emot av en målcell. Målceller har proteiner som kallas receptorer, som binder till signalmolekyler och orsakar ett svar. Signalmolekyler som binder till receptorer kallas ligander. Ligander och receptorer är specifika för varandra; en receptor binder vanligtvis endast till sin specifika ligand. Det finns dock olika typer av signalering.
9.1.1 Former av signalering
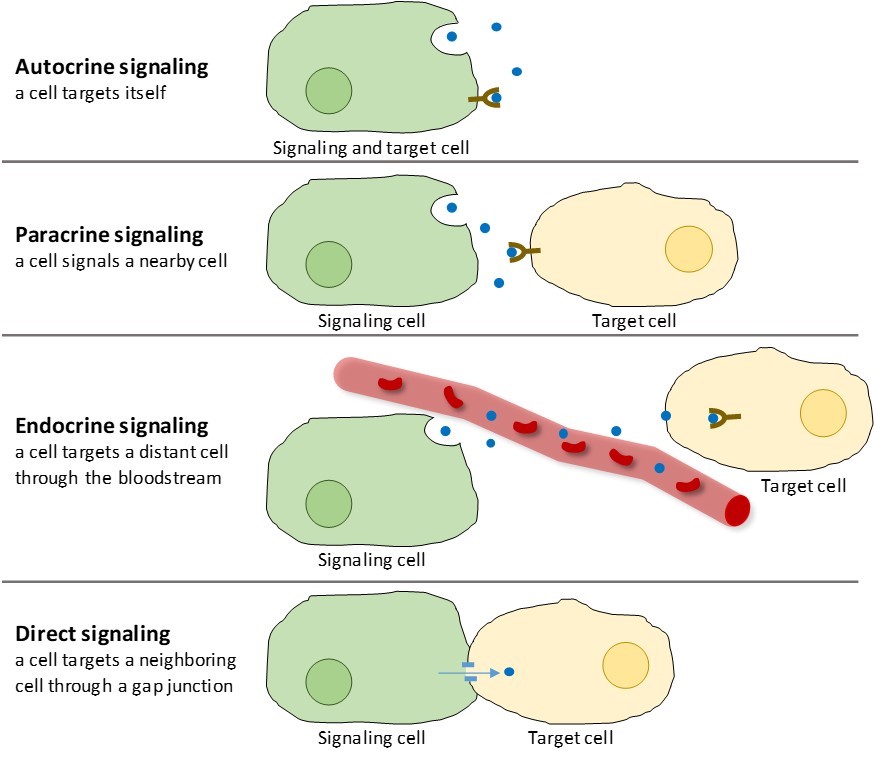
Det finns fyra kategorier av kemisk signalering som förekommer i flercelliga organismer: autokrin signalering, parakrin signalering, endokrin signalering och direkt signalering över gap junctions (figur 9.2). Den största skillnaden mellan de olika kategorierna av signalering är det avstånd som signalen färdas för att nå målcellen.
Paracrin signalering
Signaler som verkar lokalt mellan celler som befinner sig nära varandra kallas parakrina signaler. Parakrina signaler rör sig genom diffusion genom den extracellulära matrisen (figur 9.2). Dessa typer av signaler framkallar vanligtvis snabba reaktioner som endast varar en kort tid. För att hålla responsen lokaliserad brukar parakrina ligander snabbt brytas ned av enzymer eller avlägsnas av närliggande celler. När signalerna avlägsnas återupprättas koncentrationsgradienten för signalmolekylen, vilket gör att de snabbt kan diffundera genom det intracellulära utrymmet om de släpps ut igen.
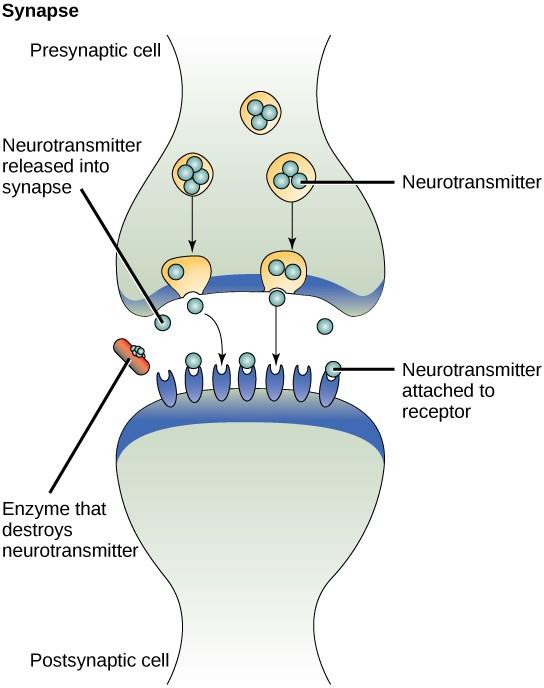
Ett exempel på parakrin signalering är överföring av signaler mellan nervceller. Det lilla utrymmet mellan nervcellerna där signalöverföring sker kallas synaps. Signalerna sprids längs nervcellerna genom snabba elektriska impulser. När dessa impulser når änden av en nervcell frigörs kemiska ligander som kallas neurotransmittorer i synapsen av den presynaptiska cellen (den cell som sänder ut signalen). Neurotransmittorerna sprids över synapsen (figur 9.3). Det lilla avståndet mellan nervcellerna gör att signalen kan färdas snabbt, vilket möjliggör en omedelbar reaktion, till exempel ”ta bort handen från spisen!”. När neurotransmittorn binder till receptorn på ytan av den postsynaptiska cellen utlöses nästa elektriska impuls. Neurotransmittorerna bryts snabbt ned eller återabsorberas av den presynaptiska cellen så att den mottagande nervcellen snabbt kan återhämta sig och vara beredd att snabbt svara på nästa synaptiska signal.
Autokrin signalering
När en cell reagerar på sin egen signalmolekyl kallas det för autokrin signalering (auto = ”själv”). Autokrin signalering förekommer ofta tillsammans med andra typer av signalering. När till exempel en parakrin signal frigörs kan den signalerande cellen svara på signalen tillsammans med sina grannar (figur 9.2).
Autokrin signalering förekommer ofta under en organisms tidiga utveckling för att se till att cellerna utvecklas till rätt vävnader. Autokrin signalering reglerar också smärtkänsla och inflammatoriska reaktioner. Vidare, om en cell infekteras med ett virus kan cellen signalera till sig själv att genomgå programmerad celldöd, vilket dödar viruset i processen.
Endokrin signalering
Signaler från avlägset belägna celler kallas endokrina signaler, och de har sitt ursprung i endokrina celler. (I kroppen finns många endokrina celler i endokrina körtlar, till exempel i sköldkörteln, hypotalamus och hypofysen). Dessa typer av signaler ger vanligtvis en långsammare reaktion men har en längrevarig effekt. De ligander som frigörs vid endokrin signalering kallas hormoner, signalmolekyler som produceras i en del av kroppen men påverkar andra kroppsregioner en bit bort (figur 9.2).
Hormoner färdas de stora avstånden mellan endokrina celler och deras målceller via blodomloppet, vilket är ett relativt långsamt sätt att förflytta sig i hela kroppen. På grund av deras transportform späds hormonerna ut och finns i låga koncentrationer när de verkar på sina målceller. Detta skiljer sig från parakrin signalering, där lokala koncentrationer av signalmolekyler kan vara mycket höga.
Direkt signalering
Gap junctions hos djur och plasmodesmata hos växter är förbindelser mellan plasmamembranen hos angränsande celler. Dessa vattenfyllda kanaler gör det möjligt för små signalmolekyler att diffundera mellan de två cellerna. Små molekyler, t.ex. kalciumjoner (Ca2+), kan röra sig mellan cellerna, men stora molekyler som proteiner och DNA kan inte ta sig igenom kanalerna. Kanalernas specificitet säkerställer att cellerna förblir oberoende men kan snabbt och enkelt överföra signaler. Direkt signalering gör det möjligt för en grupp celler att samordna sitt svar på en signal som endast en av dem kan ha fått. I växter är plasmodesmata allestädes närvarande, vilket gör hela växten till ett gigantiskt kommunikationsnätverk.
9.1.2 Typer av receptorer
Receptorer är proteinmolekyler i målcellen eller på dess yta som binder till ligander. Det finns två typer av receptorer, interna receptorer och receptorer på cellytan.
Interna receptorer
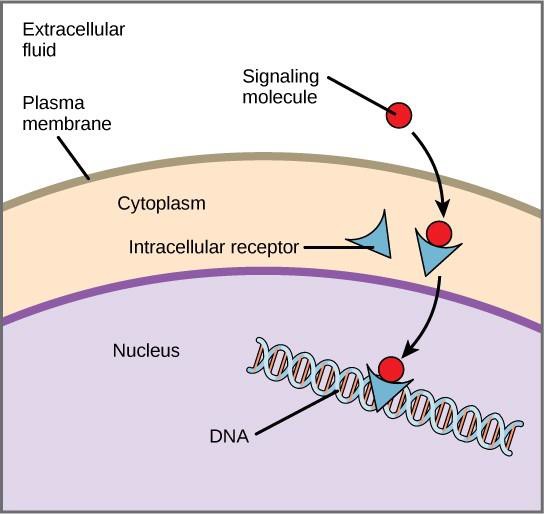
Interna receptorer, även kallade intracellulära eller cytoplasmatiska receptorer, finns i målcellernas cytoplasma och reagerar på hydrofoba ligandmolekyler som kan färdas över plasmamembranet. När de väl är inne i cellen binder många av dessa molekyler till proteiner som fungerar som regulatorer av mRNA-syntesen (transkription) för att förmedla genuttryck.
Genuttryck är den cellulära processen att omvandla informationen i en cells DNA till en sekvens av aminosyror, som slutligen bildar ett protein. När liganden binder till den interna receptorn utlöses en konformationsförändring som exponerar en DNA-bindningsplats på receptorproteinet. Ligand-receptorkomplexet rör sig in i kärnan och binder sedan till specifika reglerande regioner i kromosom-DNA:t och främjar initieringen av transkriptionen (figur 9.4). Transkription är processen att kopiera informationen i en cells DNA till en speciell form av RNA som kallas messenger RNA (mRNA); cellen använder informationen i mRNA för att koppla ihop specifika aminosyror i rätt ordning och producera ett protein. När en ligand binder till en intern receptor kan den således direkt påverka genuttrycket i målcellen.
Cellytoreceptorer
Cellytoreceptorer, även kallade transmembranreceptorer, är integrerade proteiner som binder till externa signalmolekyler. Dessa receptorer spänner över plasmamembranet och utför signaltransduktion, där en extracellulär signal omvandlas till en intercellulär signal. (Figur 9.5). Eftersom receptorproteiner på cellytan är grundläggande för cellernas normala funktion borde det inte komma som någon överraskning att en funktionsstörning i något av dessa proteiner kan få allvarliga konsekvenser. Fel i proteinstrukturerna hos vissa receptormolekyler har visat sig spela en roll vid hypertoni (högt blodtryck), astma, hjärtsjukdomar och cancer.
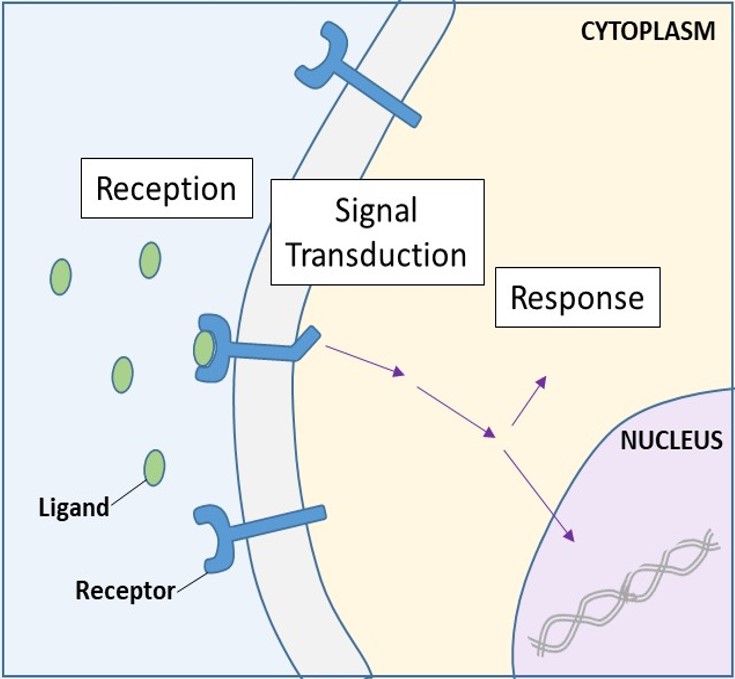
Varje receptor på cellytan har tre huvudkomponenter: en yttre ligandbindande domän, eller extracellulär domän, en hydrofob membranöverskridande region och en intracellulär domän. Receptorer på cellytan är involverade i det mesta av signaleringen i flercelliga organismer. Det finns tre allmänna kategorier av receptorer på cellytan: enzymbundna receptorer, jonkanalbundna receptorer och G-proteinbundna receptorer.
Enzymbundna receptorer är receptorer på cellytan med intracellulära domäner som är associerade med ett enzym. I vissa fall är själva receptorns intracellulära domän ett enzym. Andra enzymkopplade receptorer har en liten intracellulär domän som interagerar direkt med ett enzym. Enzymbundna receptorer har normalt stora extracellulära och intracellulära domäner, men den membranöverskridande regionen består av en enda alfa-helix i peptidsträngen.
När en ligand binder till den extracellulära domänen hos en enzymbunden receptor överförs en signal genom membranet och aktiverar enzymet. Aktiveringen av enzymet sätter igång en kedja av händelser i cellen som så småningom leder till ett svar.
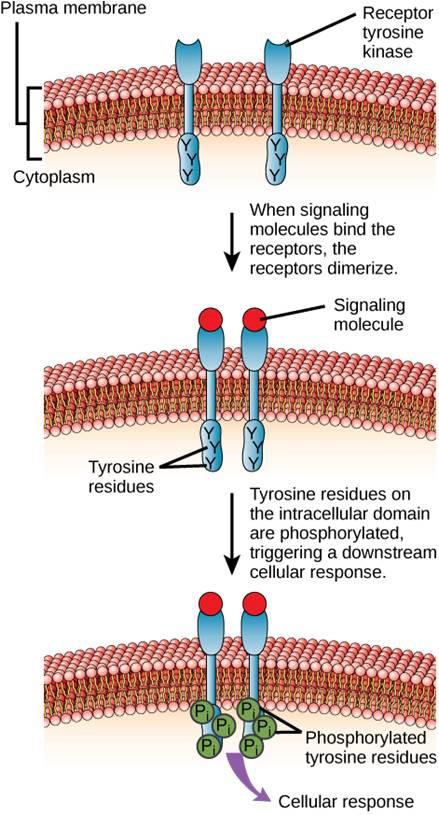
Ett exempel på en enzymkopplad receptor är tyrosinkinasreceptorn (figur 9.6). Ett kinas är ett enzym som överför fosfatgrupper från ATP till ett annat protein. Tyrosinkinasreceptorn överför fosfatgrupper till tyrosinmolekyler. Först binder signalmolekyler till den extracellulära domänen hos två närliggande tyrosinkinasreceptorer. De två närliggande receptorerna binder sedan ihop sig, eller dimeriseras. Fosfater läggs sedan till tyrosinrester på receptorernas intracellulära domän (fosforylering). De fosforylerade resterna kan sedan överföra signalen till nästa budbärare i cytoplasman.
Epidermal growth factor-receptorer är ett exempel på receptortyrosinkinaser som följer detta signaleringssätt. Defekter i ErbB-signalering i denna familj kan leda till neuromuskulära sjukdomar som multipel skleros och Alzheimers sjukdom.
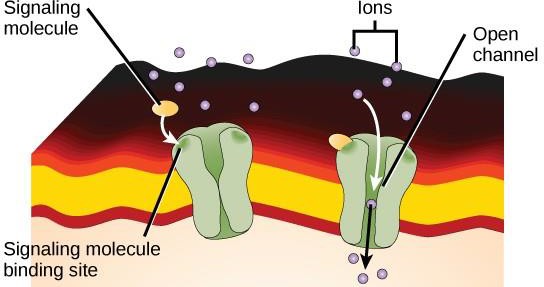
Ionkanalbundna receptorer binder till en ligand och öppnar en kanal genom membranet som tillåter specifika joner att passera. Denna typ av receptor på cellytan har en omfattande membranöverskridande region med hydrofoba aminosyror. Omvänt är de aminosyror som kantar kanalens insida hydrofila för att möjliggöra passage av joner. När en ligand binder till kanalens extracellulära region sker en konformationsförändring i proteinets struktur som gör att joner som natrium, kalcium, magnesium eller väte kan passera (figur 9.7).
G-proteinbundna receptorer binder till en ligand och aktiverar ett associerat G-protein. Det aktiverade G-proteinet interagerar sedan med ett närliggande membranprotein, som kan vara en jonkanal eller ett enzym (figur 9.8). Alla G-proteinbundna receptorer har sju transmembrandomäner, men varje receptor har en specifik extracellulär domän och G-proteinbindningsställe.
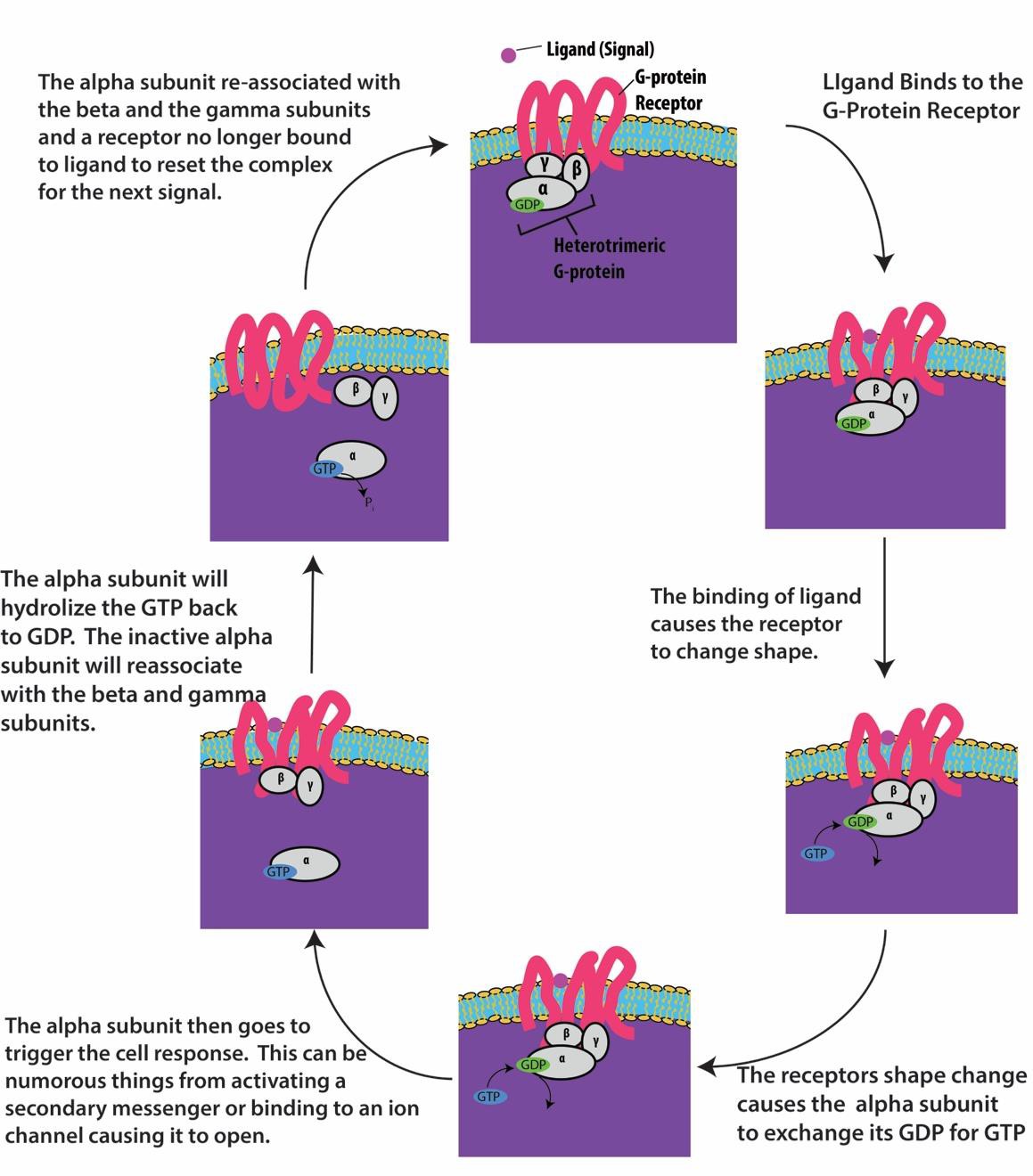
Cellsignalering med hjälp av G-proteinbundna receptorer sker som en cykel. När liganden binder till receptorn aktiverar den resulterande formförändringen G-proteinet, som frigör GDP och tar upp GTP. G-proteinets underenheter delar sig sedan i α- och βγ-underenheter. En eller båda av dessa G-proteinfragment kan aktivera andra proteiner i cellen. Efter ett tag hydrolyseras GTP på den aktiva α-underenheten av G-proteinet till GDP och βγ-underenheten avaktiveras. Underenheterna återassocieras för att bilda det inaktiva G-proteinet och cykeln börjar igen (figur 9.8).
G-proteinlänkade receptorer används i många fysiologiska processer, bland annat för syntransduktion, smak och reglering av immunsystem och inflammation.

HER2 är ett receptortyrosinkinas. I 30 procent av bröstcancer hos människor är HER2 permanent aktiverad, vilket resulterar i oreglerad celldelning. Lapatinib, ett läkemedel som används för att behandla bröstcancer, hämmar den process genom vilken receptorn fosforylerar sig själv, vilket minskar tumörtillväxten med 50 procent. Vilket av följande steg skulle hämmas av Lapatinib?
- Signalmolekylbindning, dimerisering och nedströms cellrespons
- Dimerisering och nedströms cellrespons
- Nedströms cellrespons
- Fosfatasaktivitet, dimerisering och nedströms cellrespons
9.1.3 Signalmolekyler
Ligander, som produceras av signalceller, är kemiska signaler som färdas till målceller och orsakar ett svar. De typer av molekyler som fungerar som ligander är otroligt varierande och sträcker sig från små proteiner till små joner. Ligander kategoriseras som antingen små hydrofoba ligander, som kan passera plasmamembran, eller vattenlösliga ligander, som inte kan göra det.
Små hydrofoba ligander
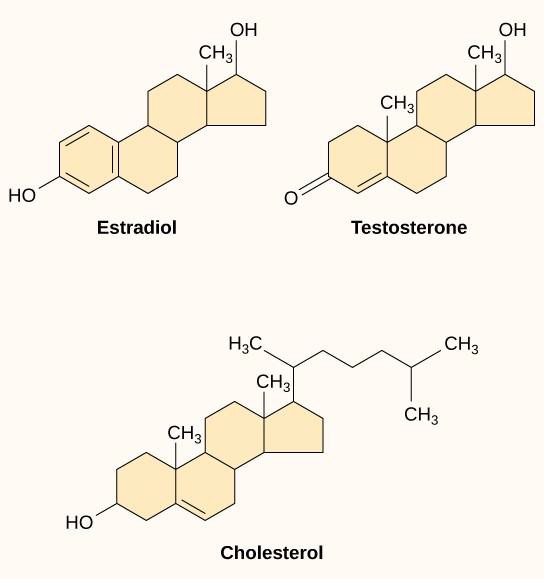
Små hydrofoba ligander, även kallade lipidlösliga ligander, kan direkt diffundera genom plasmamembranen och interagera med interna receptorer. Viktiga medlemmar av denna klass av ligander är steroidhormonerna. Steroider är lipider som har ett kolväte- skelett med fyra kondenserade ringar; olika steroider har olika funktionella grupper knutna till kolskelettet. Steroidhormoner omfattar det kvinnliga könshormonet östradiol, som är en typ av östrogen, det manliga könshormonet testosteron och kolesterol, som är en viktig strukturell komponent i biologiska membran och en föregångare till steroidhormoner (figur 9.10). Andra hydrofoba hormoner är bland annat sköldkörtelhormoner och D-vitamin. För att vara lösliga i blodet måste hydrofoba ligander binda till bärarproteiner medan de transporteras genom blodomloppet.
Vattenlösliga ligander
Då vattenlösliga ligander är polära kan de inte passera genom plasmamembranet utan hjälp. Ibland är de för stora för att överhuvudtaget kunna passera genom membranet. I stället binder de flesta vattenlösliga ligander till den extracellulära domänen hos receptorer på cellytan (se figur 9.5). Denna grupp av ligander är ganska varierande och omfattar små molekyler, peptider och proteiner.
9.2 | Signalens utbredning
I slutet av det här avsnittet kommer du att kunna:
- förklara hur bindningen av en ligand initierar signaltransduktionen i hela cellen.
- Känn igen att intracellulära signaler överförs genom fosforyleringens eller andra budbärares roll.
När en vattenlöslig ligand binder till sin receptor överförs signalen genom membranet och in i cytoplasman. Fortsättning av en signal på detta sätt kallas signaltransduktion (figur 9.5). Signaltransduktion sker endast med receptorer på cellytan eftersom interna receptorer kan ta sig in i cellen.
När en ligand binder till sin receptor sker konformationella förändringar som påverkar receptorns intracellulära domän. Dessa konformationsförändringar leder till aktivering av den intracellulära domänen eller dess associerade proteiner. I vissa fall orsakar bindning av liganden dimerisering av receptorn, vilket innebär att två receptorer binder till varandra för att bilda ett stabilt komplex som kallas dimer. En dimer är en kemisk förening som bildas när två molekyler (ofta identiska) förenas. Bindningen av receptorerna på detta sätt gör det möjligt för deras intracellulära domäner att komma i nära kontakt och aktivera varandra.
9.2.1 Signalvägar och signalförstärkning
Och även om signalmolekyler ofta finns i mycket låga koncentrationer kan de ge djupgående effekter. Efter att liganden binder till receptorn på cellytan sätter aktiveringen av receptorns intracellulära komponenter igång en kedja av händelser som kallas för en signalväg eller en signalkaskad. I en signalväg aktiverar andra budbärare, enzymer och/eller aktiverade proteiner andra proteiner eller budbärare (figur 9.11). Varje medlem i signalvägen kan aktivera tusentals av nästa medlem i signalvägen i en process som kallas signalförstärkning. Eftersom signalen förstärks i varje steg kan ett mycket stort svar genereras från en enda receptor som binder en ligand.
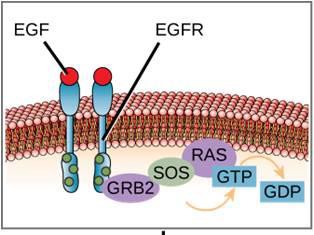
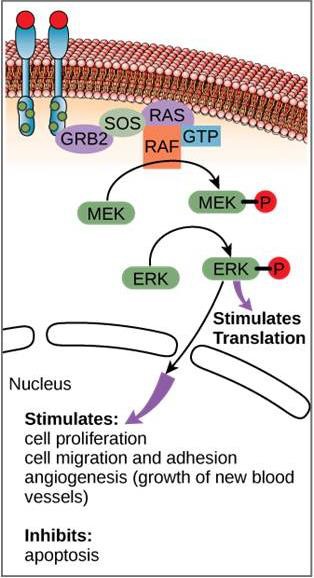
Ett exempel på en signalväg visas i figur 9.11. Epidermal tillväxtfaktor (EGF) är en signalmolekyl som är involverad i regleringen av celltillväxt, sårläkning och vävnadsreparation. Receptorn för EGF (EGFR) är ett tyrosinkinas. Ett aktiverat kinas fosforylerar och aktiverar många nedströmsmolekyler. När EGF binder till EGFR signalerar en kaskad av fosforyleringshändelser i nedströmsled till cellen att den ska växa och dela sig. Om EGFR aktiveras vid olämpliga tidpunkter kan okontrollerad celltillväxt (cancer) uppstå.
I vissa cancerformer hämmas GTPase-aktiviteten hos RAS G-proteinet. Detta innebär att RAS-proteinet inte längre kan hydrolysa GTP till GDP. Vilken effekt skulle detta ha på nedströms cellhändelser?
9.2.2 Metoder för intracellulär signalering
Induktionen av en signalväg beror på att en cellulär komponent modifieras av ett enzym. Det finns många enzymatiska modifieringar som kan ske för att aktivera nästa komponent i signalvägen. Följande är några av de vanligaste händelserna i intracellulär signalering.
Fosforylering
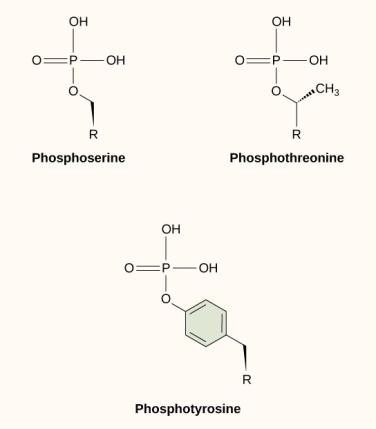
En av de vanligaste kemiska modifieringarna som sker i signalvägar är tillägget av en fosfatgrupp till en molekyl i en process som kallas fosforylering. Fosfatet kan läggas till en nukleotid, t.ex. GMP, för att bilda GDP eller GTP. Fosfater läggs också ofta till serin-, threonin- och tyrosinrester i proteiner, där de ersätter aminosyrans hydroxylgrupp (figur 9.12). Överföringen av fosfatet katalyseras av ett enzym som kallas kinas. Fosforylering kan aktivera eller inaktivera enzymer, och omvändning av fosforylering, avfosforylering, upphäver effekten.
Sekundbudbärare
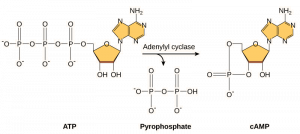
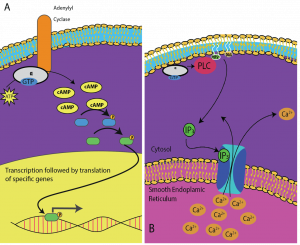
Sekundbudbärare är små molekyler som fortplantar en signal efter att den har inletts genom att signalmolekylen har bundit till receptorn. Dessa molekyler hjälper till att sprida en signal genom cytoplasman genom att ändra beteendet hos vissa cellproteiner. En andra budbärare som används av många olika celltyper är cykliskt AMP (cAMP). Cykliskt AMP syntetiseras av enzymet adenylylcyklas från ATP (figur 9.13). Den viktigaste rollen för cAMP i celler är att binda till och aktivera ett enzym som kallas cAMP-beroende kinas (A-kinas). A-kinas reglerar många viktiga metaboliska vägar: Det fosforylerar serin- och threoninrester i sina målproteiner och aktiverar dem i processen. A-kinas finns i många olika typer av celler, och målproteinerna i varje typ av cell är olika. En annan sekundär budbärare är Ca2+som kan frigöras för att översvämma cellen.
Olika celler reagerar olika på cAMP. I figur 9.14 visas alfaunderenheten från en G-proteinreceptor som aktiverar två olika typer av signalering. I den första bilden produceras cAMP av enzymet adenylatcyklas när det aktiveras av alfaunderenheten. cAMP aktiverar sedan andra proteiner som påverkar gentranskriptionen. I den andra bilden utlöser alfaunderenheten från G-proteinet en kaskad som frigör Ca2+ från det släta endoplasmatiska retikulumet. I det här fallet är Ca2+ den sekundära budbäraren som orsakar den cellulära reaktionen.
9.3 | Svar på signalen
I slutet av det här avsnittet kommer du att kunna:
- Känna igen att signalvägar styr proteinuttryck, cellulär metabolism och celltillväxt.
- Känna igen apoptosens roll i utvecklingen och upprätthållandet av en frisk organism.
Med hjälp av signaltransduktionsvägar producerar receptorer i plasmamembranet en mängd olika effekter på cellen. Inne i cellen binder ligander till sina interna receptorer, vilket gör det möjligt för dem att direkt påverka cellens DNA och proteinproducerande maskineri. Resultaten av signalvägar är extremt varierande och beror på vilken typ av cell det rör sig om samt de yttre och inre förhållandena. Ett litet urval av reaktioner beskrivs nedan.
9.3.1 Reaktioner på signalvägen
Genuttryck
Vissa signaltransduktionsvägar reglerar transkriptionen av RNA. Andra reglerar översättningen av proteiner.
Ökning av cellmetabolismen
Aktiveringen av β-adrenerga receptorer i muskelceller genom adrenalin leder till en ökning av cykliskt AMP inne i cellen. Adrenalin är ett hormon som produceras av binjuren och som förbereder kroppen för kortvariga nödsituationer.
Celltillväxt
Cellsignalvägar spelar också en viktig roll vid celldelning. Celler delar sig normalt inte om de inte stimuleras av signaler från andra celler. De ligander som främjar celltillväxt kallas tillväxtfaktorer. De flesta tillväxtfaktorer binder till receptorer på cellytan som är kopplade till tyrosinkinaser.
Celldöd
När en cell är skadad, överflödig eller potentiellt farlig för en organism kan en cell initiera en mekanism för att utlösa programmerad celldöd, eller apoptos. Apoptos gör det möjligt för en cell att dö på ett kontrollerat sätt som förhindrar att potentiellt skadliga molekyler frigörs från cellens inre. I vissa fall, t.ex. vid en virusinfektion eller okontrollerad celldelning på grund av cancer, misslyckas dock cellens normala kontroller och balanser. Extern signalering kan också initiera apoptos. De flesta normala djurceller har till exempel receptorer som interagerar med den extracellulära matrisen, ett nätverk av glykoproteiner som ger djurcellerna strukturellt stöd. Bindningen av cellulära receptorer till den extracellulära matrisen initierar en signalkaskad i cellen. Om cellen rör sig bort från den extracellulära matrisen upphör dock signalgivningen och cellen genomgår apoptos. Detta system bidrar till att förhindra att cellerna färdas genom kroppen och förökar sig okontrollerat, vilket sker med tumörceller som bildar metastaser.
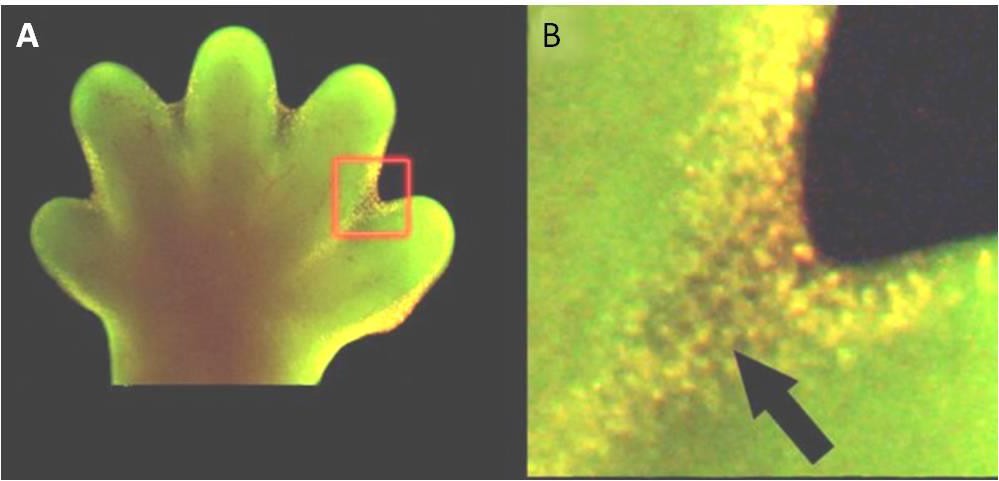
Apoptos är också viktigt för normal embryologisk utveckling. Hos ryggradsdjur omfattar till exempel de tidiga utvecklingsstadierna bildandet av nätliknande vävnad mellan enskilda fingrar och tår (figur 9.15). Under den normala utvecklingen måste dessa onödiga celler elimineras så att helt separerade fingrar och tår kan bildas. En cellsignalmekanism utlöser apoptos, vilket förstör cellerna mellan de framväxande fingrarna.
9.3.2 Avslutande av signalvägar
Den avvikande signalering som ofta ses i tumörceller är ett bevis på att avslutandet av en signal vid rätt tidpunkt kan vara lika viktigt som initieringen av en signal. En metod för att stoppa en specifik signal är att bryta ned liganden eller avlägsna den så att den inte längre kan nå sin receptor. En anledning till att hydrofoba hormoner som östrogen och testosteron utlöser långvariga händelser är att de binder till bärarproteiner. Dessa proteiner gör att de olösliga molekylerna blir lösliga i blodet, men de skyddar också hormonerna från nedbrytning av cirkulerande enzymer.
Inuti cellen reverserar många olika enzymer de cellulära modifieringar som följer av signalkaskader. Fosfataser är till exempel enzymer som tar bort den fosfatgrupp som kinaser fäster vid proteiner i en process som kallas avfosforylering. cAMP bryts ned till AMP av fosfodiesteras, vilket avslutar dess signal. På samma sätt upphävs frisättningen av kalciumlagren av Ca2+-pumpar som finns i cellmembranen.
.