Detail Description of the Experiment
- Introduction
- Materials and Methods
- Questions do dalszych przemyśleń i dyskusji
- Referencje i linki
- Narzędzia do oceny wyników nauczania studentów
- Narzędzia do oceny formatywnej tego eksperymentu
Wprowadzenie:
W tym doświadczeniu będziemy badać wpływ konkurencji o zasoby na wydajność reprodukcyjną w obrębie i pomiędzy dwoma gatunkami pasożytniczych os.
Cele laboratorium:
Po zakończeniu tego laboratorium, studenci będą w stanie…
- Opisać cykl życiowy Nasonia vitripennis i Melittobia digitata,
- Wyjaśnić możliwe interakcje między dwoma gatunkami pasożytów konkurujących o ten sam zasób żywiciela,
- Zaprojektować eksperyment w celu określenia charakteru interakcji między tymi dwoma gatunkami, gdy konkurują o wspólnego żywiciela,
- Przeprowadzić eksperyment konsensusowy w celu określenia wpływu konkurencji wewnątrzgatunkowej i międzygatunkowej na wydajność reprodukcyjną u Nasonia vitripennis i Melittobia digitata,
- Wykorzystać uzyskane dane do oszacowania parametrów modelu konkurencji Lotki-Volterry,
- Odnieść wyniki badań klasowych do zasady wykluczania konkurencji.
Zbiorowiska ekologiczne składają się z populacji wszystkich gatunków występujących w danym siedlisku. Struktura społeczności będzie określona w części przez dynamikę interakcji między gatunkami w społeczności. Interakcje między dwoma gatunkami mogą być bezpośrednie lub pośrednie (tzn. pośredniczone przez inne gatunki).
W nawet prostej naturalnej społeczności, setki różnych gatunków roślin i zwierząt wchodzą ze sobą w interakcje. Pomimo tej różnorodności, możemy jednak zidentyfikować kategorie interakcji, które mają różny wpływ na wzrost populacji (Tabela 1). Kategorie te są zdefiniowane przez kierunek wpływu na oddziałujące gatunki.
W uzupełnieniu do interakcji między gatunkami, interakcje między osobnikami w obrębie gatunku mogą być również ważne w kształtowaniu społeczności. Interakcje wewnątrzgatunkowe mogą wpływać na dynamikę populacji gatunku, która z kolei będzie wpływać na interakcje między gatunkami. Konkurencja wewnątrzgatunkowa występuje wtedy, gdy różne osobniki tego samego gatunku lub populacji konkurują o zasoby. Interakcje te mogą być zaciekłe, ponieważ osobniki wymagają tych samych ograniczonych zasobów, aby przetrwać i rozmnażać się. Gdy różne gatunki walczą o to samo pożywienie, siedlisko lub inny zasób środowiskowy, mamy do czynienia z konkurencją międzygatunkową. Te interakcje są zazwyczaj nieco mniej intensywne. Dzieje się tak dlatego, że choć wymagania dwóch gatunków mogą być podobne, nigdy nie mogą być tak zbliżone, jak w przypadku osobników tego samego gatunku.
| Tabela 1. Categories Of Direct Interactions Between Two Species In The Same Community | ||||
| Name of interaction | Type of contact | Direct effect on species #1 | Direct wpływ na gatunek #2 | Inne aspekty związku |
| Związek neutralny | Dwa gatunki są powiązane tylko pośrednio poprzez interakcje z innymi gatunkami. | 0 | 0 | Każdy gatunek ma neutralny związek z większością gatunków w swoim siedlisku. |
| Związek komensalizmu | Związek, który bezpośrednio pomaga jednemu gatunkowi, ale nie wpływa na drugi w dużym stopniu, jeśli w ogóle. | + | 0 | Komensalizm, mutualizm i pasożytnictwo są wszystkimi przypadkami symbiozy. |
| Mutualizm | Korzyści płyną w obie strony między oddziałującymi gatunkami. | + | + | Lepiej postrzegane jako dwukierunkowa eksploatacja niż jako przytulna współpraca. |
| Predation True Predators Grazers |
Predator atakuje i żywi się serią ofiar, ale nie zajmuje w nich lub na nich miejsca. | + | > | Prey generalnie umiera. W przypadku pasożytów roślina może umrzeć lub nie. |
| Pasożytnictwo Pasożyty Parazytoidy | Pasożyt odżywia się tkankami jednego lub więcej żywicieli, przebywając w nich lub na nich przez co najmniej część cyklu życia. | + | Żywiciel może umrzeć lub nie umrzeć w wyniku interakcji. | |
| Konkurencja międzygatunkowa | Wady mogą przepływać w obie strony między gatunkami, lub też lepszy konkurent może pozostać w dużej mierze nienaruszony. | Zazwyczaj mniej intensywna niż konkurencja między członkami tego samego gatunku. | ||
0 oznacza brak bezpośredniego wpływu na wzrost populacji.
+ oznacza efekt pozytywny; oznacza efekt negatywny.
Rozważmy jednak teoretyczny przypadek dwóch gatunków, które zajmują identyczną niszę. Gause (1934) badał dwa gatunki protistów, które żywiły się tymi samymi komórkami bakteryjnymi. Kiedy połączył je w jednej hodowli, jeden z nich zawsze doprowadzał drugi do wyginięcia. Od tego czasu wiele innych eksperymentów potwierdziło „prawo Gause’a”, zwane obecnie zasadą konkurencyjnego wykluczenia. Stwierdza ono, że dwa gatunki, które wykorzystują identyczne zasoby, nie mogą współistnieć w nieskończoność lub „kompletni konkurenci nie mogą współistnieć” (Hardin 1960).
Równanie dla wzrostu populacji gatunku 1 to:
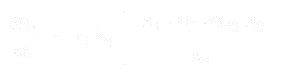
A dla gatunku 2 to:
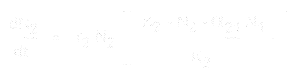
gdzie:
- * N1 i N2 to liczebności populacji gatunków 1 i 2,
* r1 i r2 to wewnętrzne tempo wzrostu dla tych gatunków,
* K1 i K2 to nośność siedliska dla każdego gatunku,
* alfa12 i alfa21 to wpływ jednego gatunku na wzrost populacji drugiego. Konkretnie, alfa12 to wpływ gatunku 2 na wzrost gatunku 1, a alfa21 to wpływ gatunku 1 na wzrost gatunku 2.
Jeśli wartości każdego równania są znane (lub mogą być oszacowane empirycznie na podstawie wyników eksperymentu), wówczas równanie może być użyte do przewidywania potencjalnego wyniku konkurencji (tj. czy gatunki mogą współistnieć, czy też jeden z nich ostatecznie wykluczy drugi). Wartości dla K1, K2, a12, i a21 są używane do wykreślenia izoklin zerowego wzrostu (tj. gdzie dN1/dt lub dN2/dt równa się zero) dla obu gatunków na tym samym wykresie, a wynikowe sumy wektorów wzrostu populacji (trajektorie) są używane do określenia wyniku konkurencji (Rysunek 1).

Rysunek 1. Przykładowe wykresy izoklin zerowego wzrostu, dla których gatunek 1 i gatunek 2 współistnieją (po lewej), a gatunek 1 konkurencyjnie wyklucza gatunek 2 (po prawej).
Model konkurencji Lotka-Volterra opisuje wynik konkurencji pomiędzy dwoma gatunkami w czasie ekologicznym. Ponieważ jeden gatunek może konkurencyjnie wykluczyć inny gatunek (Rysunek 1) w czasie ekologicznym, konkurencyjnie gorszy gatunek może zwiększyć zakres rodzajów pożywienia, które zjada, aby przetrwać. Jednakże reakcja gatunków na konkurencję międzygatunkową w czasie ewolucyjnym jest często odwrotna do tej, która ma miejsce w czasie ekologicznym. Konkurenci zazwyczaj specjalizują się w określonych typach zasobów. Ten podział zasobów, który następuje w czasie ewolucji, faktycznie skutkuje zmniejszeniem lub brakiem konkurencji pomiędzy dwoma gatunkami.
Ale nie są one szczególnie blisko spokrewnione ze sobą, historie życia dwóch gatunków pasożytniczych os, Melittobia digitata i Nasonia vitripennis, są dość podobne. Oba gatunki są zdolne do wykorzystywania tego samego żywiciela, choć w naturze używały różnych żywicieli. Melittobia są o połowę mniejsze od Nasonia, ale oba są dość małe i całkowicie nieszkodliwe dla ludzi.
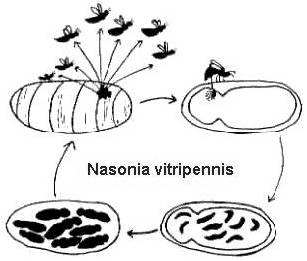
Rysunek 2. Cykl życiowy Nasonia vitripennis na poczwarce żywiciela Neobellieria bullata (rysunek Bethia King). Cykl życiowy Melittobia digitata jest taki sam, chociaż osobniki na wszystkich etapach są mniejsze.
Na świecie znanych jest około 70 000 gatunków parazytoidów (9% wszystkich owadów), ale szacunki ich liczby sięgają nawet 800 000 (Strand 2002). Spośród opisanych gatunków około 80% należy do rzędu błonkówek (Hymenoptera), podobnie jak dwa gatunki, których używasz w tym laboratorium. W naturze, żywiciele mogą być pasożytowani przez więcej niż jeden gatunek parazytoida, wszystkie konkurujące o ten sam zasób (Stand 2002). Ponieważ gatunek żywiciela może sam być pasożytem na innym gatunku, ekologiczne efekty tej konkurencji na strukturę zbiorowiska mogą być bardzo złożone. Na przykład, motyle sosny jaworowej (Neodiprion swainei Midd.), które mogą atakować, defoliacji i zabijać duże drzewostany sosny jaworowej w Ameryce Północnej, są żywicielami 11 różnych gatunków błonkówek pasożytniczych. Wydaje się, że większość z tych gatunków współistnieje, ponieważ rozdzielają one zasoby żywicielskie pomiędzy siebie, pasożytując na różnych etapach cyklu życiowego żywiciela lub różnych segmentach jego populacji (Price 1972). Jednym z członków gildii parazytoidów atakujących kokony motyli jest gatunek introdukowany (Pleolophus basizonus). Jest on lepszym konkurentem i jego obecność determinuje liczebność niektórych gatunków konkurencyjnych (Price 1970). Chociaż konkurencja może wpływać na strukturę zbiorowisk parazytoidów, to jednak przestrzenne i czasowe zróżnicowanie w zasobach żywicieli może być jeszcze ważniejsze w określaniu bogactwa gatunkowego parazytoidów, nawet przy istnieniu konkurencji międzygatunkowej wśród parazytoidów (Hawkins 2000).
Parazytoidy, których żywiciele są ważnymi szkodnikami roślin uprawnych lub drzew leśnych są czasami celowo uwalniane jako biologiczne środki kontroli populacji szkodników. Tam gdzie więcej niż jeden gatunek parazytoida atakuje tego samego żywiciela, ważne jest aby zrozumieć naturę tej konkurencji przed uwolnieniem. Jeśli różne gatunki parazytoidów są zdolne do współistnienia dzięki podziałowi zasobów, zwalczanie szkodnika może być najlepiej osiągnięte poprzez wprowadzenie niektórych lub wszystkich gatunków parazytoidów. Z drugiej strony, jeśli konkurenci ograniczają wzajemnie swoje populacje z powodu wzajemnych oddziaływań konkurencyjnych, wówczas maksymalne zwalczanie gatunków szkodników może być osiągnięte poprzez uwolnienie najbardziej efektywnego z gatunków parazytoidów (Amarasekare 2000).
Materiały i metody:
Overview of Data Collection and Analysis Methods:
Lab 1:
Kategorie interakcji omówione w tabeli 1 mogą wydawać się dość proste, gdy się o nich po prostu czyta. Ale gdybyś obserwował dwa nieznane sobie zwierzęta wchodzące w interakcję, jak zdecydowałbyś, jaką „etykietę” zastosować? Czy mógłbyś przewidzieć wynik interakcji? Jak mógłbyś sprawdzić swoje przewidywania?
Dwie parazytoidalne osy przedstawione w tym badaniu laboratoryjnym wydają się zajmować podobne nisze. Interesują nas interakcje pomiędzy tymi dwoma gatunkami. Ponadto, chcemy być w stanie określić wpływ jednego gatunku na drugi.
Każda grupa powinna:
- – przedyskutować i wymienić wszystkie możliwe kombinacje doświadczalne, które mogłyby być założone z udziałem dwóch pasożytniczych os, Melittobia digitata i Nasonia vitripennis, oraz pojedynczego żywiciela, Neobellierria,
– przewidzieć, co waszym zdaniem może być wynikiem dla każdej możliwej interakcji,
– zidentyfikuj i wymień zmienne, którymi manipulowałbyś w swoim eksperymencie,
– zidentyfikuj i wymień zmienne, które utrzymałbyś na stałym poziomie w swoim eksperymencie,
– zidentyfikuj i wymień zmienne zależne, które chciałbyś zmierzyć, aby określić ilościowo wpływ każdego gatunku na drugi.
Każda grupa podzieli się swoim projektem eksperymentu z resztą klasy. Razem, jako klasa, opracujemy wspólny projekt eksperymentu. W oparciu o uzgodniony projekt eksperymentu, każda osoba powinna założyć jedną replikę kultury dla każdego zabiegu. Patrz „Postępowanie z parazytoidami” poniżej. Będziemy zbierać dane z całej klasy do analizy.
Cotygodniowe kontrole:
W każdym tygodniu, każda osoba powinna sprawdzić swoje kultury, aby zobaczyć czy pojawiły się jakieś dorosłe osy. Powinieneś zapisać datę pierwszego pojawienia się dorosłych os dla każdej kultury. Hodowle Nasonia powinny być zamrożone 21 dni po ich założeniu. Hodowle Melittobia i mieszanych gatunków powinny być zamrożone po 42 dniach.
Lab 2 (6 tygodni po Lab 1):
Każda osoba powinna policzyć liczbę potomstwa wyprodukowanego w każdej replikowanej kulturze. Wprowadźcie swoje dane do arkusza kalkulacyjnego, aby dane dla całej klasy mogły zostać połączone. Użyjcie zebranych danych do oszacowania parametrów modelu konkurencji Lotka-Volterra i do analizy statystycznej efektów konkurencji.
W grupach przedyskutujcie, w jaki sposób dane mogą być wykorzystane do ilościowego określenia parametrów modelu konkurencji Lotka-Volterra (patrz „Ilościowe określenie modelu konkurencji Lokta-Volterra” poniżej). Przedyskutuj również, co poszczególne porównania mogą nam powiedzieć o względnym znaczeniu konkurencji wewnątrzgatunkowej i międzygatunkowej w tych dwóch gatunkach parazytoidów (patrz „Wytyczne do analizy danych” poniżej).
Postępowanie z parazytoidami:
Dorosłe osobniki obu parazytoidów, Melittobia digitata i Nasonia vitripennis, są bardzo „przyjazne dla użytkownika”. Chociaż samice posiadają normalne skrzydła i potrafią latać, nie czynią tego zbyt chętnie. Są one jednak negatywnie geotaksyczne (tzn. poruszają się w górę, z dala od grawitacji). Gdy kilka samic z hodowli zostanie wytrząśniętych na poziomą powierzchnię, a następnie przykrytych odwróconą szklaną fiolką, z łatwością wdrapią się do fiolki i po jej bokach. Gdy już masz osy w fiolce, możesz łatwo dodać poczwarkę gospodarza, a następnie szczelnie zatkać fiolkę bawełną. W ten sposób można sprawnie posługiwać się dużą liczbą osobników. Dorosłe osy mogą być również manipulowane za pomocą krótkich czyścików do rur, do których osy będą tymczasowo przylegać.
Kwantyfikacja modelu konkurencji Lotka-Volterra:
Model konkurencji Lotka-Volterra został opisany i zdefiniowany za pomocą równań we Wstępie. Jak tam zaznaczono, jeśli wartości dla każdego równania mogą być oszacowane empirycznie na podstawie wyników eksperymentu, wówczas równanie może być użyte do przewidywania potencjalnego wyniku konkurencji (tj. czy dwa gatunki będą współistnieć, czy też jeden z nich ostatecznie wykluczy drugi). Wartości dla K1, K2, alfa12, i alfa21 są używane do wykreślenia izoklin zerowego wzrostu (tj. gdzie dN1/dt lub dN2/dt równa się zero) dla obu gatunków na tym samym wykresie, a wynikowe sumy wektorów wzrostu populacji (trajektorie) są używane do określenia wyniku konkurencji.
W oparciu o nasz projekt eksperymentu musimy wyznaczyć wartości tych parametrów. Przypomnijmy, że nośność populacji to maksymalna liczba osobników, które mogą przeżyć w danym siedlisku. Dla uproszczenia w tym eksperymencie zdefiniowaliśmy siedlisko parazytoidów jako pojedynczego żywiciela. W rzeczywistości, oczywiście, siedlisko zawierałoby prawdopodobnie więcej niż jedno gniazdo babki błotnej lub puparium muchówki, a więc istniałoby wiele potencjalnie użytecznych żywicieli. Aby określić zdolność do przenoszenia dwóch gatunków, musimy znać maksymalną liczbę potomstwa danego gatunku, które może być wyprodukowane na jednym żywicielu, gdy obecny jest tylko ten gatunek. Mając to na uwadze, dane z którego zabiegu zostałyby wykorzystane do oszacowania pojemności środowiska dla Melittobia i Nasonia? (Pamiętaj, że przy nośności wszystkie zasoby żywiciela będą wykorzystane.)
Oszacowanie współczynników konkurencji (alfa12, i alfa21) jest trochę bardziej skomplikowane. Przypomnijmy, że równanie wzrostu populacji gatunku 1 ma postać:
Gdy cały zasób żywiciela zostanie wykorzystany przez pasożyty, populacja nie może już rosnąć. Innymi słowy, dN1 /dt = 0. Ten stan wystąpi, gdy K1 – N1 – alfa12*N2 = 0. Aby znaleźć alfa12, musimy ją rozwiązać (tzn. zrobić małą algebrę), a następnie podstawić wartości dla K1, N1 i N2. Powyżej opisaliśmy jak znaleźć nośność. Zakładając, że Melittobia jest „gatunkiem 1”, użyj jej nośności dla K1. Liczba potomstwa Melittobia i Nasonia produkowanego w konkurencji międzygatunkowej wynosi odpowiednio N1 i N2. Mając to na uwadze, dane z którego zabiegu byłyby użyte do oszacowania N1 i N2?
To samo podejście, które zastosowałeś do obliczenia alfa12, może być użyte do obliczenia alfa21.
Teraz, gdy obliczyłeś wszystkie wartości parametrów, możesz użyć tych wartości do wykreślenia izoklin zerowego wzrostu i przewidzieć wynik konkurencji między Melittobia i Nasonia.
Wskazówki do analizy danych:
Możemy użyć porównań między różnymi zabiegami, aby zbadać względne znaczenie konkurencji wewnątrzgatunkowej i międzygatunkowej. Najpierw należy określić, jaki typ konkurencji, wewnątrzgatunkowej czy międzygatunkowej, jeśli w ogóle, występuje w każdym zabiegu. Gdy już to zrobisz, zastanów się nad wszystkimi porównaniami pomiędzy parami obiektów. Co mówi nam każde z tych porównań? Pomocne może być stworzenie wykresu, który będzie zawierał listę porównań i ich znaczenie. Ponieważ wszystkie porównania dotyczą dwóch metod leczenia, mogą być analizowane statystycznie przy użyciu testów t.
Pytania do dalszego przemyślenia i dyskusji:
- W oparciu o wartości parametrów, które obliczyłeś dla modelu konkurencji Lotka-Volterra, jaki jest przewidywany wynik konkurencji między dwoma gatunkami? Czy przewidywany wynik został osiągnięty w każdej replikacji konkurencji międzygatunkowej? Jeśli nie, to dlaczego?
- „Prawo Gause’a” stwierdza, że konkurenci, którzy dzielą dokładnie te same zasoby w ten sam sposób, nie mogą współistnieć. Oznacza to, że gatunek, który najefektywniej wykorzystuje sporne zasoby, w końcu wyeliminuje inne w danym miejscu. Czy prawo Gause’a ma zastosowanie do interakcji między Melittobia i Nasonia? Dlaczego lub dlaczego nie?
- Gdyby te dwa gatunki używały tego samego żywiciela w naturze, jak podział zasobów mógłby pozwolić im na koegzystencję?
- W oparciu o wyniki twojego eksperymentu, dlaczego te dwa gatunki nie używają tego samego żywiciela w naturze?
- Podając wartości oszacowane dla nośności i współczynników konkurencji, przepowiedz wynik konkurencji między Melittobia i Nasonia używając modelu konkurencji Lotka-Volterra w Populus (zobacz odnośniki i linki). Czy na przewidywany wynik konkurencji ma wpływ wielkość populacji początkowej lub tempo wzrostu populacji? Jeśli tak, to w jaki sposób? Jak te wartości wpływają na czas do osiągnięcia równowagi?
- Nośność i współczynniki konkurencji są tylko szacunkowe. Jakie czynniki mogą wpłynąć na nośność i współczynniki konkurencji dla tych dwóch gatunków?
- Jeśli konkurencja międzygatunkowa występuje u tych gatunków, jak możemy określić jaki mechanizm konkurencji (interferencyjny czy eksploatacyjny) ma miejsce?
*** Uwaga: Odpowiedzi na wiele z tych pytań oraz liczne inne komentarze autora można znaleźć w „NOTATKACH DLA KIEROWNIKÓW: Comments On Questions for Further Thought” page.
Referencje i linki:
- Abe, J., Y. Kamimura, N. Kondo, and M. Shimada. 2003. Extremely female-biased sex ratio and lethal male-male combat in a parasitoid wasp, Melittobia australica (Eulophidae). Behavioural Ecology 14: 34-39.
- Amarasekare, P. 2000. Coexistence of competing parasitoids on a patchily distributed host: local vs. spatial mechanisms. Ecology 81: 1286-96.
- Cooperband M. F., R. W. Matthews, S. B. Vinson. 2003. Factors affecting the reproductive biology of Melittobia digitata (Hymenoptera: Eulophidae) and failure to meet the sex ratio predictions of Hamilton’s local mate competition theory. Entomologia Experimentalis et Applicata 109: 1-12.
- Côsoli, F. L., and S. B. Vinson. 2002. Clutch size, development and wing morph differentiation of Melittobia digitata. Entomologia Experimentalis et Applicata 102: 135-143.
- Freeman, B. E., and K. Ittyeipe. 1976. Field studies on the cumulative response of Melittobia sp. Hawaiiensis complex) (Eulophidae) to varying host densities. Journal of Animal Ecology 45: 415-423.
- Gause, G. F. 1934. The Struggle for Existence. Williams and Wilkins, Baltimore.
- Harden, G. 1960. Zasada konkurencyjnego wykluczenia. Science 131: 1292-1297.
- Hawkins, B. A. 2000. Species coexistence in parasitoid communities: does competition matter? Pages 198-213 in M. E. Hochberg and A. R. Ives (eds). Parasitoid Population Biology. Princeton University Press, Princeton, New Jersey.
- King, B. 2000. Nasonia vitripennis, osa klejnotowa. Illustrating science and biology concepts with an insect. www.bios.niu.edu/bking/nasonia.htm
- Krebs, C. J. 1994. Ecology. The Experimental Analysis of Distribution and Abundance. 4th edition. HarperCollins College Publishers, New York, NY.
- Mackauer, M., B. Bai, A. Chow, and T. Danyk. 1992. Asymmetric larval competition between two species of solitary parasitoid wasps: the influence of superparasititsm. Ecological Entomology 17: 233-236.
- Price, P. 1970. Parasitoids utilizing the same host: adaptive nature of differences in size and form. Ecology 53: 445-454.
- Price, P. 1972. Characteristics permitting coexistence among parasitoids of a sawfly in Quebec. Ecology 51: 190-195.
- Strand, M. R. 2002. The interactions between larval stage parasitoids and their hosts. Pages 129-152 in E. E. Lewis, J. F. Campbell, and M. V. K. Sukhdeo (eds). The Behavioural Ecology of Parasites. CABI Publishing, New York.
- Silva-Torres, C., and R. W. Matthews. 2003. Development of Melittobia australica Girault and M. digitata Dahms (Hymenoptera: Eulophidae) parasitizing Neobellieria bullata (Parker) (Diptera: Sarcophagidae) puparia. Neotropical Entomology 32: 645-651.
- Werren, J. H. 1983. Ewolucja stosunku płci pod lokalnym konkurencji mate w pasożytniczej osy. Evolution 37:116-124.
- Werren, J.H. 1984. Brood size and sex ratio regulation in the parasitic wasp Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae). Netherlands Journal of Zoology 34: 123-143.
W celu uzyskania informacji o historii naturalnej i siedliskach, odwiedź:
- www.wowbugs.com dla Melittobia,
www.bios.niu.edu/bking/nasonia.htm dla Nasonia,
www.rochester.edu/College/BIO/labs/WerrenLab/nasonia/ dla Nasonia.
Populus można pobrać ze strony www.cbs.umn.edu/populus.
Tools for Assessment of Student Learning Outcomes:
Assessment has been carried out in a variety of ways at the different institutions that have used this exercise. We wszystkich przypadkach studenci są oceniani na podstawie pracy naukowej napisanej przez każdego studenta indywidualnie lub przez studentów w grupie. W niektórych przypadkach studenci są oceniani na podstawie zarówno pierwszego, jak i drugiego projektu pracy.
Rubric punktacji dla papierów różni się w zależności od instruktora. Poniżej znajduje się przykładowa punktacja używana w Morehouse College dla „podsumowania wyników”, które ma wszystkie elementy pracy naukowej z wyjątkiem metod. W tej metryczce „publiczność” dotyczy wyboru odpowiedniego odbiorcy przez studenta. Oczekuje się, że studenci napiszą swój raport tak, jakby był on pracą naukową. Tak więc, odpowiednimi odbiorcami są rówieśnicy, którzy nie przeprowadzali eksperymentu, ale są obeznani z nauką. Raporty pisane do instruktora lub innych studentów w klasie nie mają odpowiedniej publiczności. „Format” to ogólna organizacja raportu w sekcjach, które mają równoległą organizację i opierają się na sobie nawzajem. Na przykład, Dyskusja powinna oceniać ustalenia przedstawione w Wynikach i umieszczać te wyniki w szerszym kontekście. Dyskusja powinna również odnosić się do hipotezy zawartej we Wstępie.
| Ocena podsumowująca wyniki (możliwe 50 punktów) | |
| Wprowadzenie i strona tytułowa (10 punktów) | _____ |
| Wyniki (10 points) | _____ |
| Discussion and Conclusions (10 points) | _____ |
| Literature Use and Citations (10 points) | _____ |
| Format, Publiczność (10 punktów) | _____ |
Na poziomie wprowadzającym, studenci zwracają wszystkie tabele wygenerowane w klasie, odpowiedzi na pytania do dyskusji i cotygodniowe notatki na temat postępów w dochodzeniu. Oprócz indywidualnych prac naukowych, studenci mogą prezentować wyniki eksperymentu w formie grupowych prac naukowych, grupowych prezentacji ustnych lub grupowych prezentacji plakatowych. Jednakże, ponieważ wszyscy uczniowie przeprowadzają ten sam eksperyment, a zatem prezentują te same wyniki, indywidualne lub grupowe prace naukowe byłyby najbardziej efektywne.
Narzędzia do oceny formatywnej tego eksperymentu:
Na kursie ekologii w Morehouse College, każdy student jest proszony o ocenę każdego ćwiczenia laboratoryjnego na 10-punktowej skali pod względem postrzeganej skuteczności każdego badania w umacnianiu ich wiedzy i zrozumienia tematów poruszanych na kursie wykładowym ekologii. Ponadto, studenci są pytani o to, które ćwiczenia były najmniej i najbardziej przyjemne oraz które najbardziej zwiększyły ich zrozumienie metody naukowej. Informacje te są wykorzystywane do wyboru i modyfikacji ćwiczeń stosowanych w kolejnych semestrach.
Na kursach ekologii na Emory University i Radford University, każdy student jest pytany, które ćwiczenia podobały mu się najbardziej, a które najmniej. Studenci czasami komentują, dlaczego ocenili ćwiczenia w taki sposób, jak to zrobili. Ponownie, informacje te są wykorzystywane do modyfikacji ćwiczeń używanych w kolejnych semestrach.
Duża dyskusja na temat Oceniania pojawia się w dziale Nauczanie na tej stronie.
.