Detail Description of the Experiment
- Introduction
- Materials and Methods
- Questions voor verder denken en discussie
- Referenties en links
- Hulpmiddelen voor de beoordeling van leerresultaten
- Hulpmiddelen voor de formatieve evaluatie van dit experiment
Inleiding:
In dit onderzoek zullen we de effecten onderzoeken van concurrentie om hulpbronnen op de voortplantingscapaciteit binnen en tussen twee soorten sluipwespen.
Doelstellingen van het laboratorium:
Aan het eind van dit lab, zullen de leerlingen in staat zijn…
- De levenscyclus van Nasonia vitripennis en Melittobia digitata beschrijven,
- De mogelijke interacties verklaren tussen twee parasietensoorten die concurreren voor dezelfde gastheerbron,
- Een experiment ontwerpen om de aard van de interactie tussen deze twee soorten te bepalen wanneer ze concurreren voor een gemeenschappelijke gastheer,
- Een consensus experiment uitvoeren om de effecten van intraspecifieke en interspecifieke competitie op de voortplantingsoutput bij Nasonia vitripennis en Melittobia digitata te bepalen,
- De resulterende gegevens gebruiken om de parameters van het Lotka-Volterra competitiemodel te schatten,
- De resultaten van het klasse-onderzoek relateren aan het principe van competitie-uitsluiting.
Ecologische gemeenschappen zijn samengesteld uit populaties van alle soorten in een habitat. De structuur van een gemeenschap wordt mede bepaald door de dynamiek van de interacties tussen de soorten in de gemeenschap. Interacties tussen twee soorten kunnen direct of indirect zijn (d.w.z. bemiddeld door andere soorten).
In zelfs een eenvoudige natuurlijke gemeenschap staan honderden verschillende soorten planten en dieren met elkaar in wisselwerking. Ondanks deze diversiteit kunnen we categorieën van interacties identificeren die verschillende effecten hebben op de populatiegroei (tabel 1). De categorieën worden gedefinieerd door de richting van de effecten op de interagerende soorten.
Naast interacties tussen soorten kunnen ook interacties tussen individuen binnen een soort van belang zijn voor de structurering van een gemeenschap. Interacties binnen een soort kunnen van invloed zijn op de populatiedynamiek van de soort, die op haar beurt weer van invloed is op interacties tussen soorten. Intraspecifieke concurrentie doet zich voor wanneer verschillende individuen van dezelfde soort of populatie met elkaar concurreren om een hulpbron. Deze interacties kunnen hevig zijn omdat de individuen dezelfde beperkte hulpbronnen nodig hebben om te overleven en zich voort te planten. Wanneer verschillende soorten wedijveren om hetzelfde voedsel, dezelfde habitat of een andere hulpbron in het milieu, is er sprake van interspecifieke concurrentie. Deze interacties zijn over het algemeen iets minder intens. Dit komt doordat de behoeften van twee soorten weliswaar vergelijkbaar kunnen zijn, maar nooit zo dicht bij elkaar kunnen liggen als bij individuen van dezelfde soort.
| Tabel 1. Categorieën van directe interacties tussen twee soorten in dezelfde gemeenschap | ||||||
| Naam van de interactie | Type contact | Direct effect op soort #1 | Direct effect op soort #2 | Direct effect op soort #2 | . effect op soort #2 | Andere aspecten van de relatie |
| Neutrale relatie | Twee soorten zijn slechts indirect met elkaar verbonden door interacties met andere soorten. | 0 | 0 | Elke soort heeft een neutrale relatie met de meeste soorten in zijn habitat. | ||
| Commensalisme | Een relatie die de ene soort direct helpt, maar de andere niet of nauwelijks beïnvloedt. | + | 0 | Commensalisme, mutualisme, en parasitisme zijn allemaal gevallen van symbiose. | ||
| Mutualisme | Er vloeien voordelen over en weer tussen de op elkaar inwerkende soorten. | + | Beter gezien als uitbuiting in twee richtingen dan als gezellige samenwerking. | |||
| Predatie Rechte Roofdieren Grazers |
Predator valt aan en voedt zich met een reeks prooien, maar gaat er niet in of op wonen. | + | Prooi sterft meestal. Bij grazers sterft de plant al dan niet. | |||
| Parasitisme Parasieten | Parasiet voedt zich met weefsels van een of meer gastheren, en verblijft daarin of daarop gedurende ten minste een deel van zijn levenscyclus. | + | Een gastheer sterft al dan niet als gevolg van de interactie. | |||
| Interspecifieke concurrentie | Nadelen kunnen beide kanten opvloeien tussen soorten, of de superieure concurrent kan grotendeels onaangetast blijven. | Generaal minder intens dan concurrentie tussen leden van dezelfde soort. | ||||
0 betekent geen direct effect op de bevolkingsgroei.
+ betekent positief effect; betekent negatief effect.
Beschouw echter het theoretische geval van twee soorten die dezelfde niche innemen. Gause (1934) bestudeerde twee protistensoorten die zich beide voedden met dezelfde bacteriecellen. Wanneer hij ze in één cultuur combineerde, dreef de ene altijd de andere tot uitsterven. Vele andere experimenten hebben sindsdien “de Wet van Gause” ondersteund, die nu het Principe van de Concurrerende Uitsluiting wordt genoemd. Volgens deze wet kunnen twee soorten die gebruik maken van identieke hulpbronnen niet onbeperkt naast elkaar bestaan of kunnen “volledige concurrenten niet naast elkaar bestaan” (Hardin 1960).
Vele experimenten hebben aangetoond dat hoe meer twee soorten in een habitat verschillen in hun gebruik van hulpbronnen, des te waarschijnlijker het is dat zij in feite naast elkaar kunnen bestaan (Krebs 1994). Zelfs twee soorten met een grote overlapping kunnen een tijdlang samenleven, hoewel concurrentiële interacties vaak de groeisnelheid van een van beide of van beide soorten onderdrukken. Na verloop van tijd kan zich een interessant verschijnsel voordoen dat “resource partitioning” wordt genoemd. Leden van elke soort kunnen zich gaan specialiseren in een onderverdeling van een categorie van vergelijkbare hulpbronnen. Bijvoorbeeld, als beide zich voeden met appels, kan de ene zich voeden met kleine groene vruchten en de andere met grotere, rijpere vruchten.
Het Lotka-Volterra-model werd ontwikkeld om ecologen in staat te stellen de mogelijke uitkomst te voorspellen wanneer twee soorten met elkaar concurreren om dezelfde hulpbronnen. In wezen tracht het model rekening te houden met het effect dat de aanwezigheid van de ene soort zal hebben op de populatiegroei van de andere soort, in vergelijking met het concurrentie-effect dat twee leden van dezelfde soort op elkaar zouden hebben.
De vergelijking voor de populatiegroei van soort 1 is:
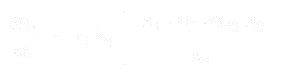
En voor soort 2 is deze:
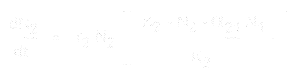
waar:
- * N1 en N2 zijn de populatiegroottes van soorten 1 en 2,
* r1 en r2 zijn de intrinsieke groeisnelheden voor deze soorten,
* K1 en K2 zijn de draagkracht van de habitat voor elke soort,
* alpha12 en alpha21 zijn de effecten van de ene soort op de populatiegroei van de andere. Meer bepaald is alpha12 het effect van soort 2 op de groei van soort 1, en alpha21 het effect van soort 1 op de groei van soort 2.
Als de waarden voor elke vergelijking bekend zijn (of empirisch kunnen worden geschat op grond van de resultaten van een experiment), kan de vergelijking worden gebruikt om de mogelijke uitkomst van een competitie te voorspellen (d.w.z. of de twee soorten naast elkaar kunnen bestaan of dat de ene uiteindelijk de andere zal uitsluiten). De waarden voor K1, K2, a12 en a21 worden gebruikt om de isoclijnen van nulgroei (d.w.z. waar dN1/dt of dN2/dt gelijk is aan nul) voor beide soorten op dezelfde grafiek uit te zetten, en de resulterende sommen van populatiegroeivectoren (trajectoriën) worden gebruikt om de uitkomst van de concurrentie te bepalen (figuur 1).

Figuur 1. Voorbeeldgrafieken van isoclines van nulgroei waarbij soort 1 en soort 2 naast elkaar bestaan (links), en waarbij soort 1 soort 2 competitief uitsluit (rechts).
Het Lotka-Volterra-competitiemodel beschrijft het resultaat van concurrentie tussen twee soorten in ecologische tijd. Omdat een soort een andere soort (figuur 1) in ecologische tijd competitief kan uitsluiten, kan de inferieure soort het scala aan voedseltypen dat hij eet vergroten om te overleven. De reactie van soorten op interspecifieke concurrentie in evolutionaire tijd is echter vaak het tegenovergestelde van wat in ecologische tijd gebeurt. Concurrenten zullen zich over het algemeen specialiseren op bepaalde typen hulpbronnen. Deze verdeling van hulpbronnen, die in de loop van de evolutie optreedt, leidt in feite tot een vermindering of het ontbreken van concurrentie tussen de twee soorten.
Hoewel zij niet bijzonder nauw aan elkaar verwant zijn, vertoont de levensloop van twee sluipwespensoorten, Melittobia digitata en Nasonia vitripennis, een grote gelijkenis. Beide soorten zijn in staat dezelfde gastheer te gebruiken, hoewel ze in de natuur verschillende gastheren gebruikten. Melittobia zijn ongeveer half zo groot als Nasonia, maar beide zijn vrij klein en volkomen ongevaarlijk voor de mens.
Hun volledige levenscycli zijn betrekkelijk kort (2-4 weken bij 25o C), en lijken ook vrij sterk op elkaar (figuur 2). De vrouwtjes leggen talrijke eitjes in de bekleding van de gastheer. De eitjes komen uit en worden larven die de gastheer verorberen, vervolgens veranderen in poppen en tenslotte metamorfoseren tot een volwassen stadium. Bij Melitobbia digitata kunnen de volwassen wijfjes normale of stompe vleugels hebben. De normaal gevleugelde volwassen dieren verspreiden zich van de gastheer om op zoek te gaan naar nieuw voedsel. De loopvlugge wijfjes leggen hun eitjes op dezelfde gastheer waaruit ze zijn voortgekomen, of verspreiden zich naar een nieuwe gastheer binnen hetzelfde nest (Freeman en Ittyeipe 1976, Côsoli en Vinson 2002).
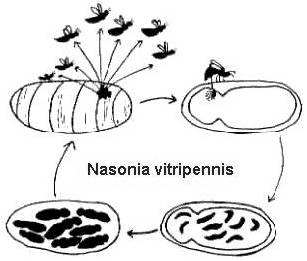
Figuur 2. De levenscyclus van Nasonia vitripennis op een Neobellieria bullata-gastheerpop (tekening door Bethia King). De levenscyclus van Melittobia digitata is dezelfde, hoewel de individuen in alle stadia kleiner zijn.
Er zijn wereldwijd ongeveer 70.000 soorten parasitoïden bekend (9% van alle insecten), maar schattingen van hun aantal lopen op tot 800.000 (Strand 2002). Van de beschreven soorten behoort ongeveer 80% tot de orde vliesvleugeligen (Hymenoptera), net als de twee soorten die je in dit practicum gebruikt. In de natuur kunnen gastheren geparasiteerd worden door meer dan één soort parasitoïde, die allemaal concurreren om dezelfde bron (Strand 2002). Aangezien de gastheersoort zelf een parasiet kan zijn op een andere soort, kunnen de ecologische effecten van die concurrentie op de gemeenschapsstructuur zeer complex zijn. Bijvoorbeeld, Swaine jack pine zaagvliegen (Neodiprion swainei Midd.), die grote opstanden van jack pines in Noord Amerika kunnen aanvallen, ontbladeren en doden, zijn gastheer voor 11 verschillende soorten parasitoïde wespen. Het blijkt dat de meeste van deze soorten naast elkaar bestaan omdat zij de gastheerbron onder elkaar verdelen door verschillende stadia van de levenscyclus van de gastheer, of verschillende segmenten van de gastheerpopulatie te parasiteren (Price 1972). Eén lid van het parasitoïdengilde dat cocons van zaagwespen aanvalt, is een geïntroduceerde soort (Pleolophus basizonus). Het is een superieure concurrent en zijn aanwezigheid bepaalt de overvloed van sommige van zijn concurrerende soorten (Price 1970). Hoewel concurrentie van invloed kan zijn op de structuur van de parasitoïdengemeenschap, is de ruimtelijke en temporele variatie in gastheerbronnen wellicht nog belangrijker voor het bepalen van de soortenrijkdom van parasitoïden, zelfs in aanwezigheid van interspecifieke concurrentie tussen parasitoïden (Hawkins 2000).
Parasitoïden waarvan de gastheren belangrijke plagen zijn op gewassen of bosbomen worden soms opzettelijk uitgezet als biologische bestrijders van plaagdierpopulaties. Wanneer meer dan één soort parasitoïde dezelfde gastheer aanvalt, is het belangrijk om de aard van die concurrentie te begrijpen alvorens tot uitzetting over te gaan. Als de verschillende parasitoïdesoorten naast elkaar kunnen bestaan door het verdelen van de hulpbronnen, kan de plaag wellicht het best worden bestreden door sommige of alle parasitoïdesoorten uit te zetten. Anderzijds, indien de concurrenten elkaars populaties beperken door hun onderlinge competitieve interacties, kan een maximale bestrijding van de plaag worden bereikt door het uitzetten van de meest efficiënte van de parasitoïdensoorten alleen (Amarasekare 2000).
Materialen en methoden:
Overzicht van gegevensverzamelings- en analysemethoden:
Lab 1:
De categorieën interacties die in tabel 1 worden besproken, kunnen vrij eenvoudig lijken als je er alleen maar over leest. Maar als je twee onbekende dieren in interactie zou observeren, hoe zou je dan beslissen welk “label” je zou toepassen? Zou je de uitkomst van de interactie kunnen voorspellen? Hoe zou je je voorspelling kunnen testen?
De twee parasitoïde wespen die in dit laboratoriumonderzoek worden gepresenteerd, lijken vergelijkbare niches in te nemen. Wij zijn geïnteresseerd in de interacties tussen de twee soorten. Bovendien willen we het effect van de ene soort op de andere kunnen kwantificeren.
Elke groep moet:
- – alle mogelijke experimentele combinaties bespreken en opnoemen met twee sluipwespen, Melittobia digitata en Nasonia vitripennis, en één gastheer, Neobellierria,
– voorspellen wat volgens jou de uitkomst zou kunnen zijn voor elke mogelijke interactie,
– bepaal en noem variabelen die je in je experiment zou manipuleren,
– bepaal en noem variabelen die je in je experiment constant zou houden,
– bepaal en noem afhankelijke variabelen die je zou willen meten om het effect van elke soort op de andere te kwantificeren.
Elke groep zal zijn experimentele opzet met de rest van de klas delen. Samen als klas ontwikkelen we een experimenteel consensusontwerp. Op basis van de consensus van de proefopzet moet iedereen voor elke behandeling een replicaatcultuur opzetten. Zie “Omgaan met Parasitoïden” hieronder. We zullen de gegevens van de hele klas samenvoegen voor analyse.
Wekelijkse controles:
Elke week moet iedereen zijn kweek controleren om te zien of er volwassen wespen zijn opgekomen. Noteer voor elke kweek de datum waarop u voor het eerst volwassen wespen hebt gezien. Nasonia-kweken moeten 21 dagen na de kweek worden ingevroren. Kweken van Melittobia en gemengde soorten moeten na 42 dagen worden ingevroren.
Lab 2 (6 weken na lab 1):
Elk persoon moet het aantal nakomelingen tellen dat in elke replicaatkweek is geproduceerd. Voer uw gegevens in een spreadsheet in, zodat de gegevens voor de klas kunnen worden samengevoegd. Gebruik de gepoolde gegevens voor het schatten van de parameters voor het Lotka-Volterra concurrentiemodel en voor statistische analyse van de effecten van concurrentie.
Bespreek in jullie groepjes hoe de gegevens kunnen worden gebruikt om de parameters van het Lotka-Volterra concurrentiemodel te kwantificeren (zie “Kwantificering van het Lokta-Volterra concurrentiemodel” hieronder). Bespreek ook wat specifieke vergelijkingen tussen behandelingen ons kunnen vertellen over het relatieve belang van intraspecifieke en interspecifieke competitie bij deze twee soorten parasitoïden (zie “Richtlijnen voor gegevensanalyse” hieronder).
Hantering van parasitoïden:
De volwassen exemplaren van beide parasitoïden, Melittobia digitata en Nasonia vitripennis, zijn zeer “gebruiksvriendelijk”. Hoewel de vrouwtjes normale vleugels hebben en kunnen vliegen, doen ze dit niet gemakkelijk. Ze zijn echter negatief geotaxisch (d.w.z. ze bewegen zich omhoog, weg van de zwaartekracht). Wanneer een paar vrouwtjes uit een kweek op een horizontaal oppervlak worden geschud en vervolgens worden afgedekt met een omgekeerd glazen flesje, zullen zij gemakkelijk in het flesje klimmen en langs de wanden omhoog. Als je eenmaal wespen in een flesje hebt, kun je er gemakkelijk een gastheerpop aan toevoegen en het flesje goed afsluiten met katoen. Grote aantallen individuen kunnen op deze manier efficiënt worden behandeld. De volwassen wespen kunnen ook worden gemanipuleerd met korte pijpenragers, waaraan de wespen zich tijdelijk zullen hechten.
Kwantificering van het Lotka-Volterra competitiemodel:
Het Lotka-Volterra-competitiemodel werd in de inleiding beschreven en gedefinieerd met vergelijkingen. Zoals daar werd opgemerkt, kan, als de waarden voor elke vergelijking empirisch kunnen worden geschat op grond van de resultaten van een experiment, de vergelijking worden gebruikt om de mogelijke uitkomst van een competitie te voorspellen (d.w.z. of de twee soorten naast elkaar zullen bestaan of dat de ene uiteindelijk de andere zal uitsluiten). De waarden voor K1, K2, alpha12 en alpha21 worden gebruikt om de isoclines van nulgroei (d.w.z. waar dN1/dt of dN2/dt gelijk is aan nul) voor beide soorten op dezelfde grafiek uit te zetten, en de resulterende sommen van populatiegroeivectoren (trajectoriën) worden gebruikt om de uitkomst van de concurrentie te bepalen.
Op basis van onze experimentele opzet moeten we de waarden van deze parameters bepalen. Bedenk dat de draagkracht van een populatie het maximale aantal individuen is dat in een habitat kan overleven. Voor de eenvoud hebben we in dit experiment de habitat van de parasitoïden gedefinieerd als een enkele gastheer. In werkelijkheid zal een habitat natuurlijk meer dan één nest of poparium van een bromvlieg bevatten, en zullen er dus vele mogelijke gastheren zijn. Om de draagkracht van de twee soorten te bepalen, moeten we weten hoeveel nakomelingen van een bepaalde soort maximaal kunnen worden geproduceerd op één gastheer wanneer alleen die soort aanwezig is. Met dit in gedachten, zouden de gegevens van welke behandeling worden gebruikt om de draagkracht van Melittobia en Nasonia te schatten? (Bedenk dat bij de draagkracht alle gastheerbronnen worden gebruikt.)
Het schatten van de concurrentiecoëfficiënten (alpha12, en alpha21) is iets ingewikkelder. Herinner u dat de vergelijking voor de populatiegroei van soort 1 luidt:
Wanneer alle gastheerbronnen door de parasitoïden zijn gebruikt, kan een populatie niet meer groeien. Met andere woorden, dN1 /dt = 0. Deze voorwaarde doet zich voor wanneer K1 – N1 – alpha12*N2 = 0. Om alpha12 te vinden, moeten we het oplossen (d.w.z. een beetje algebra doen) en vervolgens de waarden voor K1, N1, en N2 substitueren. Hierboven hebben we beschreven hoe we de draagkracht kunnen vinden. Ervan uitgaande dat Melittobia “soort 1” is, gebruik dan zijn draagkracht voor K1. Het aantal nakomelingen van Melittobia en Nasonia dat bij interspecifieke competitie wordt geproduceerd, is respectievelijk N1 en N2. Met dit in gedachten, gegevens van welke behandeling zou worden gebruikt om N1 en N2 te schatten?
Dezelfde aanpak die u gebruikte om alpha12 te berekenen, kan worden gebruikt om alpha21 te berekenen.
Nu u alle parameterwaarden hebt berekend, kunt u deze waarden gebruiken om de nulgroei-isoclijnen uit te zetten en de uitkomst van de concurrentie tussen Melittobia en Nasonia te voorspellen.
Richtlijnen voor gegevensanalyse:
Wij kunnen vergelijkingen tussen verschillende behandelingen gebruiken om het relatieve belang van intraspecifieke en interspecifieke concurrentie te onderzoeken. Stel eerst vast welk type concurrentie, intraspecifiek of interspecifiek, bij elke behandeling eventueel optreedt. Als je dit gedaan hebt, denk dan na over alle vergelijkingen tussen de behandelingsparen. Wat zegt elk van de vergelijkingen ons? Het kan nuttig zijn een schema te maken met een lijst van de vergelijkingen en wat ze betekenen. Aangezien alle vergelijkingen twee behandelingen betreffen, kunnen ze statistisch worden geanalyseerd met behulp van t-tests.
Vragen voor verder denken en discussie:
- Baseer je op de parameterwaarden die je voor het Lotka-Volterra concurrentiemodel hebt berekend, wat is dan de voorspelde uitkomst van de concurrentie tussen de twee soorten? Werd de voorspelde uitkomst in elke replicatie van interspecifieke competitie bereikt? Zo niet, waarom niet?
- De Wet van Gause stelt dat concurrenten die precies dezelfde hulpbronnen op dezelfde manier delen, niet naast elkaar kunnen bestaan. Dit betekent dat de soort die het meest efficiënt gebruikmaakt van de betwiste hulpbron, de andere op die plaats uiteindelijk zal elimineren. Lijkt de Wet van Gause van toepassing te zijn op de interactie tussen Melittobia en Nasonia? Waarom wel of niet?
- Als deze twee soorten in de natuur dezelfde gastheer zouden gebruiken, hoe zou de verdeling van hulpbronnen hen dan in staat stellen naast elkaar te bestaan?
- Baseert u zich op de resultaten van uw experiment, waarom gebruiken de twee soorten in de natuur dan niet dezelfde gastheer?
- Voorspel, gegeven de geschatte waarden voor de draagkracht en de concurrentiecoëfficiënten, de uitkomst van de concurrentie tussen Melittobia en Nasonia met behulp van het Lotka-Volterra-competitiemodel in Populus (zie Referenties en links). Wordt de voorspelde uitkomst van de competitie beïnvloed door de initiële populatiegrootte of populatiegroeisnelheid? Zo ja, hoe? Hoe wordt de tijd die nodig is om een evenwicht te bereiken door deze waarden beïnvloed?
- De draagvermogens en concurrentiecoëfficiënten zijn slechts schattingen. Welke factoren kunnen van invloed zijn op de draagkracht en de concurrentiecoëfficiënt voor deze twee soorten?
- Als er bij deze soorten sprake is van interspecifieke concurrentie, hoe kunnen we dan bepalen welk mechanisme van concurrentie (interferentie of uitbuiting) optreedt?
*** Opmerking: Antwoorden op veel van deze vragen en tal van andere opmerkingen van de bijdragende auteur zijn te vinden in de “NOTES TO FACULTY: Commentaren op vragen om verder over na te denken”.
Referenties en links:
- Abe, J., Y. Kamimura, N. Kondo, and M. Shimada. 2003. Extremely female-biased sex ratio and lethal male-male combat in a parasitoid wasp, Melittobia australica (Eulophidae). Behavioural Ecology 14: 34-39.
- Amarasekare, P. 2000. Coëxistentie van concurrerende parasitoïden op een fragmentarisch verspreide gastheer: lokale vs. ruimtelijke mechanismen. Ecology 81: 1286-96.
- Cooperband M. F., R. W. Matthews, S. B. Vinson. 2003. Factors affecting the reproductive biology of Melittobia digitata (Hymenoptera: Eulophidae) and failure to meet the sex ratio predictions of Hamilton’s local mate competition theory. Entomologia Experimentalis et Applicata 109: 1-12.
- Côsoli, F. L., en S. B. Vinson. 2002. Legselgrootte, ontwikkeling en vleugelmorfdifferentiatie bij Melittobia digitata. Entomologia Experimentalis et Applicata 102: 135-143.
- Freeman, B. E., and K. Ittyeipe. 1976. Field studies on the cumulative response of Melittobia sp. Hawaiiensis complex) (Eulophidae) to varying host densities. Journal of Animal Ecology 45: 415-423.
- Gause, G. F. 1934. The Struggle for Existence. Williams and Wilkins, Baltimore.
- Harden, G. 1960. Het principe van uitsluiting door concurrentie. Science 131: 1292-1297.
- Hawkins, B. A. 2000. Species coexistence in parasitoid communities: does competition matter? Pages 198-213 in M. E. Hochberg and A. R. Ives (eds). Parasitoïden Populatie Biologie. Princeton University Press, Princeton, New Jersey.
- King, B. 2000. Nasonia vitripennis, de juweelwesp. Wetenschappelijke en biologische concepten illustreren met een insect. www.bios.niu.edu/bking/nasonia.htm
- Krebs, C. J. 1994. Ecologie. The Experimental Analysis of Distribution and Abundance. 4e editie. HarperCollins College Publishers, New York, NY.
- Mackauer, M., B. Bai, A. Chow, and T. Danyk. 1992. Asymmetrische larvale competitie tussen twee soorten solitaire parasitoïde wespen: de invloed van superparasitieten. Ecological Entomology 17: 233-236.
- Price, P. 1970. Parasitoids utilizing the same host: adaptive nature of differences in size and form. Ecology 53: 445-454.
- Price, P. 1972. Kenmerken die coëxistentie tussen parasitoïden van een zaagwesp in Quebec mogelijk maken. Ecology 51: 190-195.
- Strand, M. R. 2002. De interacties tussen parasitoïden in het larvale stadium en hun gastheren. Pages 129-152 in E. E. Lewis, J. F. Campbell, and M. V. K. Sukhdeo (eds). The Behavioural Ecology of Parasites. CABI Publishing, New York.
- Silva-Torres, C., and R. W. Matthews. 2003. Ontwikkeling van Melittobia australica Girault en M. digitata Dahms (Hymenoptera: Eulophidae) die parasiteren op Neobellieria bullata (Parker) (Diptera: Sarcophagidae) poparia. Neotropical Entomology 32: 645-651.
- Werren, J. H. 1983. Sex ratio evolution under local mate competition in a parasitic wasp. Evolution 37:116-124.
- Werren, J.H. 1984. Regeling van broedselgrootte en sexratio bij de sluipwesp Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae). Nederlands Tijdschrift voor Dierkunde 34: 123-143.
Voor informatie over natuurlijke historie en habitats, zie:
- www.wowbugs.com voor Melittobia,
www.bios.niu.edu/bking/nasonia.htm voor Nasonia,
www.rochester.edu/College/BIO/labs/WerrenLab/nasonia/ voor Nasonia.
Populus kan worden gedownload van www.cbs.umn.edu/populus.
Tools for Assessment of Student Learning Outcomes:
De evaluatie is op verschillende manieren uitgevoerd in de verschillende instellingen die deze methode hebben gebruikt. In alle gevallen worden de studenten geëvalueerd op basis van een wetenschappelijk werkstuk dat door elke student afzonderlijk of door studenten in een groep wordt geschreven. In sommige gevallen worden zowel de eerste als de tweede versie van een werkstuk beoordeeld.
De quotering voor de werkstukken varieert per docent. Hieronder staat een voorbeeld van een beoordelingsschema dat aan Morehouse College wordt gebruikt voor een “samenvatting van de resultaten”, die alle onderdelen van een wetenschappelijk werkstuk bevat behalve de methoden. In deze evaluatierubriek heeft “publiek” betrekking op de keuze van het geschikte publiek door de student. Van de leerlingen wordt verwacht dat ze hun verslag schrijven alsof het een wetenschappelijk artikel is. Het juiste publiek is dus dat van medestudenten die het experiment niet hebben uitgevoerd, maar die wel wetenschappelijk onderlegd zijn. Verslagen die aan de docent of aan andere studenten in de klas worden geschreven, hebben niet het juiste publiek. “Formaat” is de algemene organisatie van het verslag in secties die parallel lopen en op elkaar voortbouwen. Bijvoorbeeld, de Discussie moet de bevindingen in de Resultaten evalueren en deze resultaten in een groter verband plaatsen. De Discussie moet ook ingaan op de hypothese die in de Inleiding is geformuleerd.
| Evaluatie van de resultaten (50 punten mogelijk) | |
| Inleiding en titelpagina (10 punten) | _____ |
| Resultaten (10 punten) | _____ |
| Discussie en conclusies (10 punten) | _____ |
| Literatuurgebruik en citaten (10 punten) | _____ |
| Opmaak, Publiek (10 punten) | _____ |
Op het inleidende niveau leveren de studenten alle klassikaal gegenereerde tabellen in, de antwoorden op de Discussievragen, en wekelijkse notities over de voortgang van het onderzoek. Naast individuele wetenschappelijke werkstukken zouden de leerlingen de resultaten van het experiment kunnen presenteren in de vorm van wetenschappelijke werkstukken voor groepen, mondelinge presentaties voor groepen of posterpresentaties voor groepen. Maar omdat alle leerlingen hetzelfde experiment uitvoeren en dus dezelfde resultaten presenteren, zouden individuele of groepswerkstukken het meest effectief zijn.
Hulpmiddelen voor de formatieve evaluatie van dit experiment:
In de cursus ecologie aan het Morehouse College wordt elke student gevraagd om elke laboratoriumoefening op een schaal van 10 punten te beoordelen op het punt van de waargenomen doeltreffendheid van elk onderzoek bij het versterken van hun kennis en begrip van de onderwerpen die in de cursus ecologie aan de orde zijn gekomen. Bovendien wordt de studenten gevraagd welke oefeningen het minst en welke het meest plezierig waren en welke hun inzicht in de wetenschappelijke methode het meest hebben vergroot. Deze informatie wordt gebruikt om de oefeningen voor de volgende semesters te kiezen en aan te passen.
In de ecologiecursussen aan de Emory University en de Radford University wordt elke student gevraagd welke oefeningen hij het leukst en welke hij het minst leuk vond. Studenten geven soms commentaar op de vraag waarom zij de oefeningen zo beoordeelden. Ook deze informatie wordt gebruikt om de oefeningen in de volgende semesters aan te passen.
Een uitgebreide discussie over evaluatie is te vinden in het gedeelte over lesgeven op deze site.