Detailed Description of the Experiment
- Introduction
- Materials and Methods
- Questions továbbgondolásra és megvitatásra
- Hivatkozások és hivatkozások
- A tanulói tanulási eredmények értékelésének eszközei
- A kísérlet formatív értékelésének eszközei
Bevezetés:
Ezzel a vizsgálattal az erőforrásokért folytatott versengésnek a szaporodási teljesítményre gyakorolt hatását vizsgáljuk két parazitoid darázsfajon belül és között.
Laborcélkitűzések:
A laboratórium végén a tanulók képesek lesznek…
- Leírni a Nasonia vitripennis és a Melittobia digitata életciklusát,
- Megmagyarázni az ugyanazon gazdaszervezet erőforrásáért versengő két parazita faj közötti lehetséges kölcsönhatásokat,
- Kísérletet tervezni a két faj közötti kölcsönhatás jellegének meghatározására, amikor egy közös gazdaszervezetért versengenek,
- Végezzen konszenzuskísérletet a Nasonia vitripennis és a Melittobia digitata fajon belüli és fajközi versengés szaporodási teljesítményre gyakorolt hatásainak meghatározására,
- A kapott adatok felhasználásával becsülje meg a Lotka-Volterra-féle versengési modell paramétereit,
- Kapcsolja össze az osztálykutatás eredményeit a versengés kizárásának elvével.
Az ökológiai közösségek egy élőhely összes fajának populációiból állnak. Egy közösség szerkezetét részben a közösségben élő fajok közötti kölcsönhatások dinamikája határozza meg. Két faj közötti kölcsönhatás lehet közvetlen vagy közvetett (azaz más fajokon keresztül közvetített).
Még egy egyszerű természetes közösségben is több száz különböző növény- és állatfaj lép kölcsönhatásba egymással. E sokféleség ellenére azonban azonosíthatjuk a kölcsönhatások kategóriáit, amelyek különböző hatással vannak a populáció növekedésére (1. táblázat). A kategóriákat a kölcsönhatásban lévő fajokra gyakorolt hatások iránya határozza meg.
A fajok közötti kölcsönhatások mellett a fajon belüli egyedek közötti kölcsönhatások is fontosak lehetnek egy közösség strukturálásában. A fajon belüli kölcsönhatások befolyásolhatják a faj populációdinamikáját, ami viszont befolyásolja a fajok közötti kölcsönhatásokat. A fajon belüli versengés akkor következik be, amikor ugyanazon faj vagy populáció különböző egyedei versengenek egy erőforrásért. Ezek a kölcsönhatások hevesek lehetnek, mivel az egyedeknek ugyanazokra a korlátozott erőforrásokra van szükségük a túléléshez és a szaporodáshoz. Amikor a különböző fajok ugyanazért a táplálékért, élőhelyért vagy más környezeti erőforrásért versengenek, ezt nevezzük interspecifikus versengésnek. Ezek a kölcsönhatások általában kevésbé intenzívek. Ennek oka, hogy bár két faj igényei hasonlóak lehetnek, de soha nem lehetnek olyan közel egymáshoz, mint az azonos faj egyedei esetében.
| 1. táblázat. Az azonos közösségben élő két faj közötti közvetlen kölcsönhatások kategóriái | |||||
| A kölcsönhatás neve | A kapcsolat típusa | Közvetlen hatás az 1. fajra | közvetlen. hatás a 2. fajra | A kapcsolat egyéb aspektusai | |
| Semleges kapcsolat | Két faj csak közvetve, más fajokkal való kölcsönhatáson keresztül kapcsolódik. | 0 | 0 | Minden faj semleges kapcsolatban áll az élőhelyén élő legtöbb fajjal. | |
| Kompromisszum | Kapcsolat, amely közvetlenül segíti az egyik fajt, de a másik fajra nincs nagy hatással, ha egyáltalán van. | + | 0 | A kommenzalizmus, a mutualizmus és a parazitizmus mind a szimbiózis esetei. | |
| Mutualizmus | A hasznok mindkét irányba áramlanak a kölcsönhatásban lévő fajok között. | + | + | Jobb kétirányú kihasználásnak tekinteni, mint meghitt együttműködésnek. | |
| Ragadozó Az igazi ragadozók A legelők |
A ragadozó megtámad egy sor zsákmányt és táplálkozik belőlük, de nem telepszik meg bennük vagy rajtuk. | + | A zsákmány általában elpusztul. A legelők esetében a növény elpusztulhat, de az is lehet, hogy nem pusztul el. | ||
| Parazitizmus Paraziták Parazitoidok | A parazita egy vagy több gazdatest szöveteivel táplálkozik, és életciklusának legalább egy részében bennük vagy rajtuk tartózkodik. | + | A gazdatest a kölcsönhatás következtében elpusztulhat, de az is lehet, hogy nem. | ||
| Specifikuson belüli verseny | Hátrányok mindkét irányba áramolhatnak a fajok között, vagy a felsőbbrendű versenytárs nagyrészt érintetlen maradhat. | Általában kevésbé intenzív, mint az azonos faj tagjai közötti verseny. | |||
0 azt jelenti, hogy nincs közvetlen hatása a populáció növekedésére.
+ pozitív hatást jelent; negatív hatást jelent.
Vizsgáljuk meg azonban azt az elméleti esetet, amikor két faj azonos fülkét foglal el. Gause (1934) két olyan protista fajt vizsgált, amelyek mindketten ugyanazokkal a baktériumsejtekkel táplálkoztak. Amikor egyetlen kultúrában egyesítette őket, az egyik mindig kipusztulásba kergette a másikat. Azóta számos más kísérlet is alátámasztotta a “Gause-törvényt”, amelyet ma a kompetitív kizárás elvének neveznek. Ez kimondja, hogy két azonos erőforrásokat használó faj nem létezhet egymás mellett a végtelenségig, vagy “teljes versenytársak nem létezhetnek egymás mellett” (Hardin 1960).
Számos kísérlet bizonyította, hogy minél inkább különbözik két faj egy élőhelyen az erőforrás-felhasználásukban, annál valószínűbb, hogy valójában együtt tudnak létezni (Krebs 1994). Még két, egymást nagymértékben átfedő faj is élhet együtt egy ideig, bár a kompetitív kölcsönhatások gyakran elnyomják az egyik vagy mindkét faj növekedési ütemét. Idővel az erőforrások felosztásának nevezett érdekes jelenség léphet fel. Az egyes fajok tagjai specializálódhatnak a hasonló erőforrások valamelyik kategóriájának felosztására. Például, ha mindketten almával táplálkoznak, az egyikük a kis zöld gyümölcsökkel, a másikuk pedig a nagyobb, érettebb gyümölcsökkel táplálkozik.
A Lotka-Volterra-modellt azért fejlesztették ki, hogy az ökológusok megjósolhassák a lehetséges kimeneteleket, amikor két faj verseng ugyanazokért az erőforrásokért. A modell alapvetően azt a hatást próbálja figyelembe venni, amelyet az egyik faj jelenléte gyakorol a másik faj populációjának növekedésére, ahhoz a versenyhatáshoz képest, amelyet ugyanazon faj két tagja gyakorolna egymásra.
Az 1. faj populációnövekedésének egyenlete:
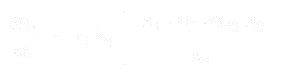
A 2. faj esetében pedig:
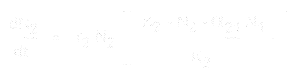
ahol:
- * N1 és N2 az 1. és 2. faj populációjának mérete,
* r1 és r2 e fajok belső növekedési rátája,
* K1 és K2 az élőhely eltartóképessége mindkét faj számára,
* alfa12 és alfa21 az egyik faj hatása a másik faj populációnövekedésére. Pontosabban, alpha12 a 2. faj hatása az 1. faj növekedésére, és alpha21 az 1. faj hatása a 2. faj növekedésére.
Ha az egyes egyenletek értékei ismertek (vagy empirikusan becsülhetők egy kísérlet eredményeiből), akkor az egyenlet felhasználható a versengés lehetséges kimenetelének előrejelzésére (azaz, hogy képesek-e együtt létezni, vagy az egyik végül kizárja a másikat). A K1, K2, a12 és a21 értékeit arra használjuk, hogy a két faj számára a zérus növekedés izoklináit (azaz ahol a dN1/dt vagy a dN2/dt egyenlő nullával) ugyanazon a grafikonon ábrázoljuk, és az így kapott populációnövekedési vektorok (pályák) összegét használjuk a verseny kimenetelének meghatározására (1. ábra).

1. ábra. Példa grafikonok a nulla növekedésű izoklinákról, amelyek esetében az 1. és a 2. faj együtt él (balra), és az 1. faj kompetitív módon kizárja a 2. fajt (jobbra).
A Lotka-Volterra versenymodell leírja két faj közötti verseny kimenetelét az ökológiai időben. Mivel az egyik faj az ökológiai időben kompetitív módon kizárhat egy másik fajt (1. ábra), a kompetitív módon alulmaradó faj a túlélés érdekében növelheti az általa elfogyasztott tápláléktípusok körét. A fajok evolúciós időben a fajok közötti versengésre adott válasza azonban gyakran az ellenkezője annak, ami az ökológiai időben történik. A versenytársak általában bizonyos erőforrástípusokra specializálódnak. Ez az evolúciós időben bekövetkező erőforrás-felosztás valójában a két faj közötti verseny csökkenését vagy hiányát eredményezi.
Bár nem állnak különösebben közeli rokonságban egymással, két parazitoid darázsfaj, a Melittobia digitata és a Nasonia vitripennis élettörténete meglehetősen hasonló. Mindkét faj képes ugyanazt a gazdát használni, bár a természetben különböző gazdákat használtak. A Melittobia körülbelül fele akkora, mint a Nasonia, de mindkettő meglehetősen kicsi és az emberre teljesen ártalmatlan.
A teljes életciklusuk viszonylag rövid (2-4 hét 25o C-on), és szintén meglehetősen hasonló (2. ábra). A nőstények számos petét raknak le a gazdatest borításán keresztül. A petékből lárvák kelnek ki, amelyek elfogyasztják a gazdaszervezetet, majd bábokká alakulnak át, és végül kifejlett stádiumba metamorfizálódnak. A Melitobbia digitata esetében a kifejlett nőstényeknek normális vagy csonka szárnyaik lehetnek. A normál szárnyú kifejlett egyedek szétszélednek a gazdaszervezetből, hogy új táplálékforrásokat keressenek. A röpképtelen nőstények ugyanarra a gazdaszervezetre rakják le petéiket, amelyből kikerültek, vagy ugyanazon a fészken belül új gazdaszervezetre szóródnak szét (Freeman és Ittyeipe 1976, Côsoli és Vinson 2002).
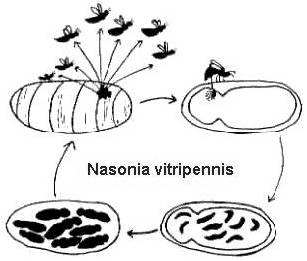
2. ábra. A Nasonia vitripennis életciklusa egy Neobellieria bullata gazdabábon (rajz: Bethia King). A Melittobia digitata életciklusa ugyanez, bár az egyedek minden stádiumban kisebbek.
A parazitoidáknak világszerte mintegy 70 000 faja ismert (az összes rovar 9%-a), de a becslések szerint számuk 800 000-re tehető (Strand 2002). A leírt fajok mintegy 80%-a a Hymenoptera rend tagja, mint ahogy az a két faj is, amelyet ebben a laborban használsz. A természetben a gazdaszervezeteket egynél több parazitoid faj is parazitálhatja, amelyek mind ugyanazon erőforrásért versengenek (Stand 2002). Mivel a gazdafaj maga is lehet egy másik faj parazitája, ennek a versengésnek a közösség szerkezetére gyakorolt ökológiai hatásai nagyon összetettek lehetnek. Például a Swaine-fenyőfűrészlepke (Neodiprion swainei Midd.), amely Észak-Amerikában nagy fenyőállományokat képes megtámadni, lombtalanítani és elpusztítani, 11 különböző parazitoid darázsfaj gazdája. Úgy tűnik, hogy e fajok többsége azért létezik egymás mellett, mert a gazdaszervezet életciklusának különböző szakaszaiban vagy a gazdapopuláció különböző szegmenseiben élősködve felosztják egymás között a gazdaszervezet erőforrásait (Price 1972). A fűrészlegyek gubóit megtámadó parazitoid céh egyik tagja egy betelepített faj (Pleolophus basizonus). Ez egy kiváló konkurens, és jelenléte meghatározza néhány konkurens faj abundanciáját (Price 1970). Bár a versengés befolyásolhatja a parazitoidok közösségének szerkezetét, a gazdaszervezetek erőforrásainak térbeli és időbeli változása még fontosabb lehet a parazitoid fajgazdagság meghatározásában, még a parazitoidok közötti fajközi versengés jelenlétében is (Hawkins 2000).
Azokat a parazitoidákat, amelyek gazdanövényei fontos kártevők a termesztett növényeken vagy erdei fákon, néha szándékosan szabadítják ki biológiai védekezésként a kártevő populációkra. Ha egynél több parazitoidafaj támadja meg ugyanazt a gazdaszervezetet, fontos, hogy a kibocsátások előtt megértsük a versengés természetét. Ha a különböző parazitoid fajok az erőforrások felosztása révén képesek egymás mellett létezni, a kártevő gazdaszervezet elleni védekezés a parazitoid fajok egy részének vagy mindegyikének betelepítésével érhető el a legjobban. Másrészt, ha a konkurensek versenykapcsolataik miatt korlátozzák egymás populációit, akkor a kártevőfajok maximális kontrollját a parazitoid fajok közül a leghatékonyabb egyedek szabadon engedésével lehet elérni (Amarasekare 2000).
Anyagok és módszerek:
Áttekintés az adatgyűjtési és elemzési módszerekről:
Labor 1:
Az 1. táblázatban tárgyalt kölcsönhatási kategóriák meglehetősen egyszerűnek tűnhetnek, ha egyszerűen csak olvasunk róluk. De ha két ismeretlen állat kölcsönhatását kellene megfigyelnünk, hogyan döntenénk el, hogy milyen “címkét” alkalmazzunk? Meg tudnád jósolni a kölcsönhatás kimenetelét? Hogyan tudnád tesztelni a jóslatodat?
Az ebben a laboratóriumi vizsgálatban bemutatott két parazitoid darázs látszólag hasonló fülkéket foglal el. A két faj közötti kölcsönhatások érdekelnek bennünket. Ezenkívül szeretnénk számszerűsíteni az egyik fajnak a másikra gyakorolt hatását.
Mindegyik csoportnak:
- – megvitatni és felsorolni az összes lehetséges kísérleti kombinációt, amelyet két parazita darázs, a Melittobia digitata és a Nasonia vitripennis, valamint egyetlen gazdanövény, a Neobellierria bevonásával lehetne létrehozni,
– megjósolni, hogy szerintetek mi lehet az eredménye az egyes lehetséges kölcsönhatásoknak,
– azonosítsa és sorolja fel azokat a változókat, amelyeket a kísérletben manipulálna,
– azonosítsa és sorolja fel azokat a változókat, amelyeket a kísérletben állandónak tartana,
– azonosítsa és sorolja fel azokat a függő változókat, amelyeket mérni szeretne, hogy számszerűsítse az egyes fajok egymásra gyakorolt hatását.
Minden csoport ossza meg kísérleti tervét az osztály többi tagjával. Osztályként közösen kidolgozunk egy konszenzusos kísérleti tervet. A konszenzusos kísérleti terv alapján mindenki állítson fel egy-egy ismétlődő tenyészetet minden kezeléshez. Lásd alább a “Parazitoidok kezelése” című részt. Az egész osztály adatait összesítjük az elemzéshez.
Heti ellenőrzések:
Minden héten minden személynek ellenőriznie kell a tenyészeteit, hogy keltek-e ki kifejlett darazsak. Minden kultúra esetében fel kell jegyeznie azt a dátumot, amikor először látott kifejlett egyedeket. A Nasonia-kultúrákat 21 nappal a létrehozásuk után le kell fagyasztani. A Melittobia- és vegyes fajú tenyészeteket 42 nap után kell lefagyasztani.
2. laboratórium (6 héttel az 1. laboratórium után):
Minden személynek meg kell számolnia az egyes replikátumkultúrákban termelt utódok számát. Írja be az adatait egy táblázatba, hogy az osztály adatai összesíthetők legyenek. Használja az összevont adatokat a Lotka-Volterra versenymodell paramétereinek becsléséhez és a verseny hatásainak statisztikai elemzéséhez.
A csoportjaitokban beszéljétek meg, hogyan lehet az adatokat felhasználni a Lotka-Volterra versenymodell paramétereinek számszerűsítésére (lásd alább a “A Lokta-Volterra versenymodell számszerűsítése” című részt). Vitassa meg azt is, hogy az egyes kezelések összehasonlítása mit mondhat el nekünk a fajon belüli és fajközi verseny relatív fontosságáról e két parazitoidafaj esetében (lásd az alábbi “Irányelvek az adatelemzéshez” című részt).
A parazitoidok kezelése:
Mindkét parazitoid, a Melittobia digitata és a Nasonia vitripennis felnőtt példányai nagyon “felhasználóbarátok”. Bár a nőstények normális szárnyakkal rendelkeznek és képesek repülni, ezt nem szívesen teszik. Viszont negatívan geotaxikusak (azaz felfelé, a gravitációtól távolodva mozognak). Ha néhány nőstényt egy tenyészetből vízszintes felületre rázunk ki, majd egy megfordított üvegpalackkal lefedjük őket, készségesen bemásznak az üvegbe és felkapaszkodnak az oldalán. Ha már vannak darazsak az üvegben, könnyen hozzáadhatunk egy gazdabábot, majd az üvegbe vattával szorosan betömhetjük. Nagyszámú egyedet lehet így hatékonyan kezelni. A kifejlett darazsak rövid csőtisztítókkal is manipulálhatók, amelyekre a darazsak ideiglenesen rátapadnak.
A Lotka-Volterra versenymodell számszerűsítése:
A Lotka-Volterra versenymodell leírása és egyenletekkel való meghatározása a Bevezetésben történt. Mint ott megjegyeztük, ha az egyes egyenletek értékei empirikusan megbecsülhetők egy kísérlet eredményeiből, akkor az egyenlet felhasználható a versengés lehetséges kimenetelének előrejelzésére (azaz, hogy a két faj együtt fog-e élni, vagy az egyik végül kizárja a másikat). A K1, K2, alfa12 és alfa21 értékeit arra használjuk, hogy a nulla növekedés izoklináit (azaz ahol a dN1/dt vagy a dN2/dt egyenlő nullával) mindkét fajra ugyanazon a grafikonon ábrázoljuk, és az így kapott populációnövekedési vektorok (pályák) összegét használjuk a verseny kimenetelének meghatározására.
Kísérleti tervünk alapján meg kell határoznunk ezen paraméterek értékeit. Emlékezzünk vissza, hogy egy populáció teherbíró képessége az a maximális egyedszám, amely egy élőhelyen túlélhet. Az egyszerűség kedvéért ebben a kísérletben a parazitoidák élőhelyét egyetlen gazdaszervezetként határoztuk meg. A valóságban persze egy élőhelyen valószínűleg egynél több iszaplepkefészek vagy lepkebáb található, és így sok potenciálisan hasznosítható gazda van. A két faj teherbíró képességének meghatározásához tudnunk kell, hogy egy adott fajból legfeljebb hány utódot tudnak létrehozni egyetlen gazdaszervezeten, ha csak az adott faj van jelen. Ezt szem előtt tartva, melyik kezelésből származó adatokat használnánk a Melittobia és a Nasonia hordozó kapacitásának becsléséhez? (Ne feledjük, hogy a hordozókapacitásnál az összes gazdaszervezet erőforrása felhasználásra kerül.)
A versengési együtthatók (alfa12, és alfa21) becslése egy kicsit bonyolultabb. Emlékezzünk vissza, hogy az 1. faj populációnövekedésének egyenlete:
Amikor a parazitoidok az összes gazdaszervezeti erőforrást felhasználják, akkor a populáció már nem tud növekedni. Más szóval, dN1 /dt = 0. Ez az állapot akkor következik be, ha K1 – N1 – alpha12*N2 = 0. Az alpha12 megtalálásához meg kell oldanunk (azaz egy kis algebrát kell végeznünk), majd be kell helyettesítenünk a K1, N1 és N2 értékeit. Fentebb leírtuk, hogyan találjuk meg a teherbíró képességeket. Feltételezve, hogy a Melittobia az “1. faj”, használjuk a hordozóképességét K1-re. A fajok közötti versenyben keletkező Melittobia- és Nasonia-utódok száma N1, illetve N2. Ezt szem előtt tartva, melyik kezelés adatait használnánk az N1 és N2 becsléséhez?
Az alfa12 kiszámításához ugyanazt a megközelítést használhatjuk, mint amit az alfa12 kiszámításához használtunk, az alfa21 kiszámításához is.
Most, hogy kiszámította az összes paraméterértéket, ezeket az értékeket felhasználhatja a nulla növekedési izoklinák ábrázolásához és a Melittobia és a Nasonia közötti verseny kimenetelének előrejelzéséhez.
Az adatelemzésre vonatkozó irányelvek:
A különböző kezelések közötti összehasonlítások segítségével feltárhatjuk az intraspecifikus és interspecifikus versengés relatív fontosságát. Először is azonosítsuk, hogy az egyes kezelésekben milyen típusú – intraspecifikus vagy interspecifikus – versengés zajlik, ha van ilyen. Miután ezt megtette, gondolja végig a kezelési párok közötti összes összehasonlítást. Mit árulnak el az egyes összehasonlítások? Hasznos lehet egy táblázatot készíteni, amely felsorolja az összehasonlításokat és azok jelentését. Mivel az összes összehasonlítás két kezelést érint, t-próbák segítségével statisztikailag elemezhetőek.
Kérdések továbbgondolásra és megvitatásra:
- A Lotka-Volterra versenymodellhez kiszámított paraméterértékek alapján mi a két faj közötti verseny előre jelzett eredménye? A fajok közötti versengés minden replikációjában megvalósult-e a megjósolt eredmény? Ha nem, miért nem?
- A “Gause-törvény” kimondja, hogy a pontosan ugyanazokon az erőforrásokon azonos módon osztozó versenytársak nem létezhetnek egymás mellett. Ez azt jelenti, hogy az a faj, amelyik a leghatékonyabban használja a vitatott erőforrást, előbb-utóbb ki fogja iktatni a másikat az adott helyen. Úgy tűnik, hogy Gause törvénye érvényes a Melittobia és a Nasonia közötti kölcsönhatásra? Miért vagy miért nem?
- Ha ez a két faj ugyanazt a gazdatestet használná a természetben, az erőforrások felosztása hogyan tenné lehetővé az együttélést?
- A kísérleted eredményei alapján miért nem használja a két faj ugyanazt a gazdatestet a természetben?
- A hordozóképesség és a versengési együtthatók becsült értékeinek ismeretében jósolja meg a Melittobia és a Nasonia közötti versengés kimenetelét a Lotka-Volterra versengési modell segítségével a Populusban (lásd a hivatkozások és linkek között). Befolyásolja-e a versengés megjósolt kimenetelét a kezdeti populációméret vagy a populáció növekedési üteme? Ha igen, hogyan? Hogyan befolyásolják ezek az értékek az egyensúly eléréséhez szükséges időt?
- A hordozó kapacitások és a versengési együtthatók csak becslések. Milyen tényezők befolyásolhatják a hordozókapacitásokat és a versengési együtthatókat e két faj esetében?
- Ha a fajok közötti versengés előfordul ezeknél a fajoknál, hogyan határozhatjuk meg, hogy milyen versengési mechanizmus (interferencia vagy kizsákmányolás) zajlik?
*** Megjegyzés: A válaszok számos ilyen kérdésre és a szerző számos egyéb megjegyzése megtalálható a “MEGJEGYZÉSEK A TUDOMÁNYOSOKNAK” című fejezetben: Megjegyzések a továbbgondolásra váró kérdésekhez” oldalon.
Hivatkozások és linkek:
- Abe, J., Y. Kamimura, N. Kondo és M. Shimada. 2003. Szélsőségesen nőstényes nemi arány és halálos hím-hím harc egy parazitoid darázsban, Melittobia australica (Eulophidae). Behavioural Ecology 14: 34-39.
- Amarasekare, P. 2000. Versengő parazitoidok koegzisztenciája egy foltosan eloszló gazdaszervezeten: lokális vs. térbeli mechanizmusok. Ecology 81: 1286-96.
- Cooperband M. F., R. W. Matthews, S. B. Vinson. 2003. A Melittobia digitata (Hymenoptera: Eulophidae) szaporodásbiológiáját befolyásoló tényezők és a Hamilton-féle lokális párverseny elméletének nemi arány előrejelzéseinek nem teljesülése. Entomologia Experimentalis et Applicata 109: 1-12.
- Côsoli, F. L. és S. B. Vinson. 2002. A Melittobia digitata fészekaljmérete, fejlődése és szárnymorfológiai differenciálódása. Entomologia Experimentalis et Applicata 102: 135-143.
- Freeman, B. E., and K. Ittyeipe. 1976. Terepi vizsgálatok a Melittobia sp. Hawaiiensis complex) (Eulophidae) (Eulophidae) különböző gazdasűrűségekre adott kumulatív válaszáról. Journal of Animal Ecology 45: 415-423.
- Gause, G. F. 1934. A létért folytatott küzdelem. Williams and Wilkins, Baltimore.
- Harden, G. 1960. A kompetitív kizárás elve. Science 131: 1292-1297.
- Hawkins, B. A. 2000. Fajok együttélése parazitoid közösségekben: számít-e a versengés? Oldalak 198-213 in M. E. Hochberg és A. R. Ives (szerk.). Parasitoid Population Biology. Princeton University Press, Princeton, New Jersey.
- King, B. 2000. Nasonia vitripennis, az ékszerdarázs. A természettudományos és biológiai fogalmak szemléltetése egy rovarral. www.bios.niu.edu/bking/nasonia.htm
- Krebs, C. J. 1994. Ökológia. Az elterjedés és a bőség kísérleti elemzése. Negyedik kiadás. HarperCollins College Publishers, New York, NY.
- Mackauer, M., B. Bai, A. Chow és T. Danyk. 1992. Aszimmetrikus lárvapárviadal két szoliter parazita darázsfaj között: a szuperparaziták hatása. Ecological Entomology 17: 233-236.
- Price, P. 1970. Ugyanazt a gazdaszervezetet használó parazitoidok: a méret- és alakbeli különbségek adaptív jellege. Ecology 53: 445-454.
- Price, P. 1972. Egy quebeci fűrészlepke parazitoidjai közötti együttélést lehetővé tevő jellemzők. Ecology 51: 190-195.
- Strand, M. R. 2002. A lárvastádiumú parazitoidok és gazdáik közötti kölcsönhatások. Oldalak 129-152 in E. E. Lewis, J. F. Campbell és M. V. K. Sukhdeo (szerk.). The Behavioural Ecology of Parasites. CABI Publishing, New York.
- Silva-Torres, C., and R. W. Matthews. 2003. A Neobellieria bullata (Parker) (Diptera: Sarcophagidae) bábjait parazitáló Melittobia australica Girault és M. digitata Dahms (Hymenoptera: Eulophidae) fejlődése. Neotropical Entomology 32: 645-651.
- Werren, J. H. 1983. A nemi arány evolúciója helyi párválasztási versenyben egy parazita darázsban. Evolution 37:116-124.
- Werren, J. H. 1984. Brood size and sex ratio regulation in the parasitic darázs Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae). Netherlands Journal of Zoology 34: 123-143.
A természetrajzról és az élőhelyekről szóló információkért látogasson el a következő oldalra:
- www.wowbugs.com a Melittobia,
www.bios.niu.edu/bking/nasonia.htm a Nasonia,
www.rochester.edu/College/BIO/labs/WerrenLab/nasonia/ a Nasonia.
Populus letölthető a www.cbs.umn.edu/populus oldalról.
Tools for Assessment of Student Learning Outcomes:
Az értékelést a különböző intézményekben, amelyek ezt a gyakorlatot alkalmazták, különböző módon végezték. Minden esetben a hallgatók értékelése egy tudományos dolgozat alapján történik, amelyet minden hallgató egyénileg vagy a hallgatók csoportosan írnak. Egyes esetekben a hallgatókat a dolgozat első és második vázlata alapján értékelték.
A dolgozatok pontozási rubrikája oktatótól függően változik. Az alábbiakban egy példa a Morehouse College-ban használt pontozási rubrikára egy “eredményösszefoglaló” esetében, amely a módszerek kivételével a tudományos dolgozat minden összetevőjét tartalmazza. Ebben az értékelési rubrikában a “közönség” a hallgató által választott megfelelő közönségre vonatkozik. A diákoktól elvárják, hogy úgy írják meg jelentésüket, mintha az egy tudományos dolgozat lenne. A megfelelő közönség tehát olyan kortársakból áll, akik nem végezték el a kísérletet, de tudományosan jártasak. Az oktatónak vagy az osztály többi diákjának írt jelentéseknek nincs megfelelő közönsége. A “formátum” a jelentés általános szervezése olyan szakaszokban, amelyek párhuzamos szervezésűek és egymásra épülnek. Például a Megbeszélésnek értékelnie kell az Eredményekben közölt eredményeket, és ezeket az eredményeket tágabb kontextusba kell helyezni. A Megbeszélésnek a Bevezetésben megfogalmazott hipotézisre is ki kell térnie.
| Eredmények összefoglaló értékelése (50 pont lehetséges) | |
| Bevezetés és címoldal (10 pont) | _____ |
| Eredmények (10 pont). pont) | _____ |
| Diskurzus és következtetések (10 pont) | _____ |
| Irodalomhasználat és idézetek (10 pont) | _____ |
| Formátum, Közönség (10 pont) | _____ |
A bevezető szinten a tanulók leadják az összes, az osztály által készített táblázatot, a vitakérdésekre adott válaszokat, valamint heti jegyzeteket a vizsgálat menetéről. Az egyéni tudományos dolgozatok mellett a tanulók a kísérlet eredményeit csoportos tudományos dolgozatok, csoportos szóbeli előadások vagy csoportos poszterbemutatók formájában is bemutathatják. Mivel azonban minden tanuló ugyanazt a kísérletet végzi, és ezért ugyanazokat az eredményeket mutatja be, az egyéni vagy csoportos tudományos dolgozatok lennének a leghatékonyabbak.
A kísérlet formatív értékelésének eszközei:
A Morehouse College ökológia kurzusán minden hallgatót arra kérnek, hogy egy 10 pontos skálán értékelje az egyes laboratóriumi gyakorlatokat abból a szempontból, hogy az egyes vizsgálatok milyen hatékonyságot érzékeltek az ökológia előadáson tárgyalt témák ismereteinek és megértésének megerősítésében. Ezenkívül a hallgatókat megkérdezik, hogy mely gyakorlatok voltak a legkevésbé és a legélvezetesebbek, és melyek növelték leginkább a tudományos módszer megértését. Ezt az információt felhasználják a következő félévekben használt gyakorlatok kiválasztásához és módosításához.
Az Emory University és a Radford University ökológia kurzusain minden hallgatót megkérdeznek arról, hogy melyik gyakorlat tetszett nekik a legjobban és melyik a legkevésbé. A hallgatók néha megjegyzéseket fűznek ahhoz, hogy miért értékelték a gyakorlatokat úgy, ahogyan értékelték. Ezt az információt is felhasználják a következő félévekben használt gyakorlatok módosításához.
Az értékelésről szóló részletes értekezés a honlap Tanítás című részében található.