
ここに例を入力する
- 9.1 信号分子と細胞レセプター
- 9.1 信号分子と細胞レセプター
- 9.2 シグナルの伝播
- 9.3 シグナルに対する応答
あなたとあなたの周りの人々がコミュニケーションできなかったら、どんな生活になるか想像してみてください。 自分の希望を他人に伝えることはできないし、自分の環境について詳しく知るために質問することもできないでしょう。 社会組織は、その社会を構成する個人間のコミュニケーションに依存しています。
人と同様に、個々の細胞にとっても、環境および互いに相互作用できることが重要です。 これは、細胞が池の中で単独で成長している場合でも、より大きな生物を形成する多くの細胞の1つである場合でも同じである。 多細胞生物では、細胞は常に化学的メッセージを送受信し、他の器官、組織、および細胞の作用を調整する。 メッセージを迅速かつ効率的に送信する能力により、細胞はその機能を調整し微調整することができる。 大きな生物では細胞間通信の必要性は明らかだが、単細胞生物でさえも互いに通信している。 酵母の細胞は、交配を助けるために互いに信号を送り合っている。 また、ある種のバクテリアは、バイオフィルムと呼ばれる大きな複合体を形成するために、あるいは、競合する生物を排除するために毒素を生産するために、互いに協調して行動している。 化学信号による細胞のコミュニケーション能力は、単細胞から生まれ、多細胞生物の進化に不可欠であった。 効率的でエラーのないコミュニケーションは、すべての生命にとって不可欠です。
- 9.1|Signaling Molecules and Cellular Receptors
- 9.1.1 シグナル伝達の形態
- パラクリンシグナル伝達
- 内分泌シグナル
- ダイレクトシグナリング
- 9.1.2 受容体の種類
- 内部受容体
- 細胞表面受容体
- 9.1.3 シグナル分子
- Small Hydrophobic Ligands
- 水溶性リガンド
- 9.2 | シグナルの伝播
- 9.2.1 シグナル伝達経路とシグナル増幅
- 9.2.2 細胞内シグナリングの方法
- リン酸化
- Second Messenger
- 9.3 | シグナルへの応答
- 9.3.1 シグナル伝達経路に対する応答
- Gene Expression
- 細胞代謝の増加
- 細胞成長
- 細胞死
- 9.3.2 シグナル伝達経路の終了
9.1|Signaling Molecules and Cellular Receptors
このセクションの終わりまでに、あなたは以下のことができるようになります:
- 多細胞生物に見られる4種類のシグナル伝達を記述します。
- 内部受容体と細胞表面受容体を比較する。
- リガンドの化学的性質とその作用機序の関係を認識する。
生きた細胞の世界では2種類のコミュニケーションが存在する。 細胞間のコミュニケーションは細胞間シグナリングと呼ばれ、細胞内のコミュニケーションは細胞内シグナリングと呼ばれます。 この違いを簡単に覚えるには、接頭語のinter-は「間」(州間を横断する州間高速道路)、intra-は「内側」(IVは静脈内または「静脈内」)を意味することを理解すればよい。
化学信号は信号伝達細胞から放出され、標的細胞で受信される。 標的細胞には受容体と呼ばれるタンパク質があり、これがシグナル伝達分子と結合して反応を引き起こす。 受容体に結合するシグナル伝達分子はリガンドと呼ばれる。 リガンドと受容体は互いに特異的であり、受容体は通常、その特定のリガンドにのみ結合する。
9.1.1 シグナル伝達の形態
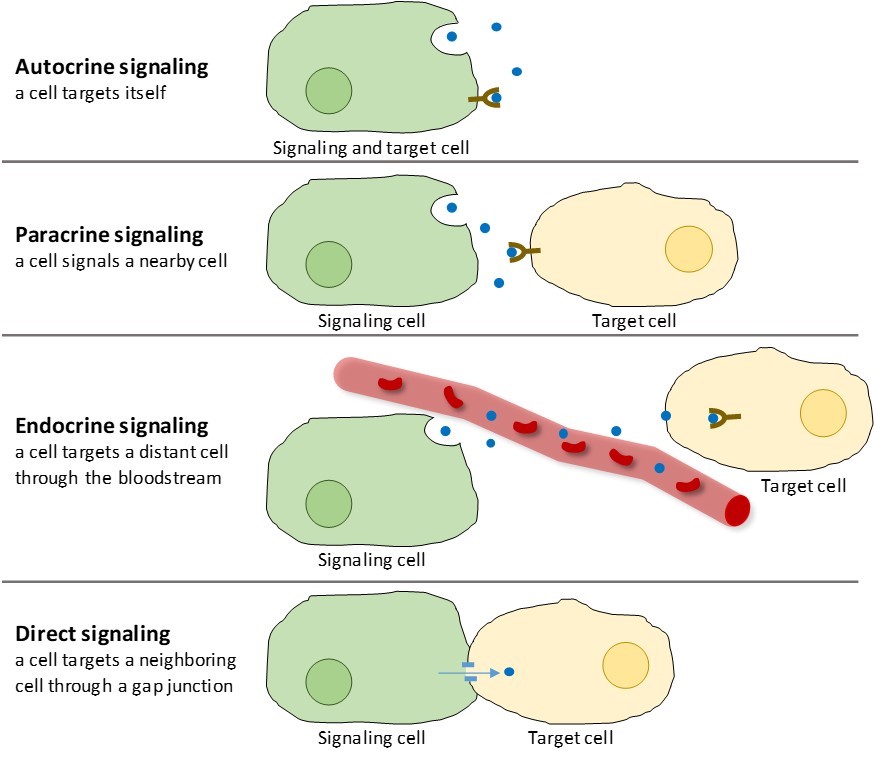
多細胞生物に見られる化学シグナル伝達には、オートクラインシグナル、パラクラインシグナル、内分泌シグナル、ギャップジャンクションを介した直接シグナルの4種類がある(図9.2)。
パラクリンシグナル伝達
近接している細胞間で局所的に作用するシグナルはパラクリンシグナルと呼ばれる。 パラクリンシグナルは細胞外マトリックス中を拡散することで移動する(図9.2)。 この種のシグナルは、通常、短時間しか続かない素早い反応を引き起こす。 反応を局所に留めるため、パラクリンリガンドは通常、酵素によって速やかに分解されるか、隣接する細胞によって除去される。 シグナルを除去することにより、シグナル分子の濃度勾配が再び確立され、再び放出された場合、細胞内空間を速やかに拡散することができる。
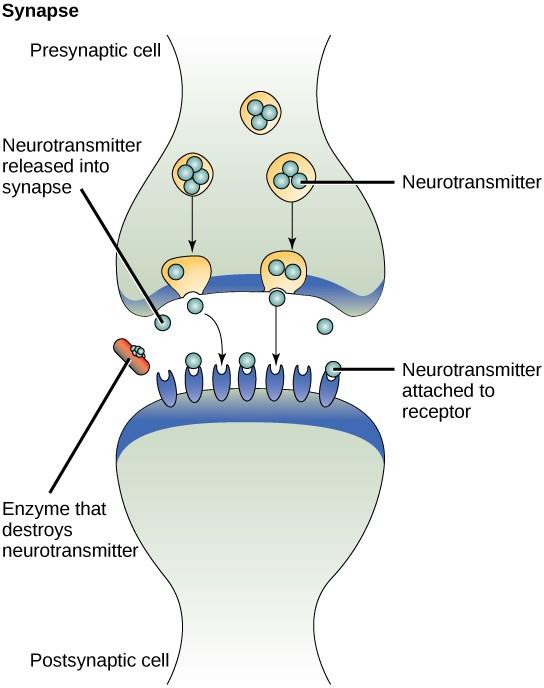
パラクラインシグナルの一例は、神経細胞間のシグナルの伝達である。 信号伝達が行われる神経細胞間の小さな空間は、シナプスと呼ばれる。 シグナルは、高速で移動する電気的インパルスによって神経細胞に沿って伝搬される。 このインパルスが神経細胞の末端に到達すると、シナプス前細胞(信号を発する細胞)から神経伝達物質という化学的リガンドがシナプスに放出されます。 神経伝達物質はシナプスを横切って拡散する(図9.3)。 神経細胞間の距離が小さいため、信号は素早く伝達され、「ストーブから手を離せ!」といった反応が即座に得られる。 神経伝達物質がシナプス後細胞の表面にある受容体と結合すると、次の電気インパルスが発 生する。 神経伝達物質は速やかに分解されるか、シナプス前細胞に再吸収されるので、受信側の神経細胞は速やかに回復し、次のシナプス信号に対して速やかに反応できるように準備される。 オートクラインシグナルはしばしば他のタイプのシグナル伝達と一緒に発生する。 例えば、パラクラインシグナルが放出されると、シグナル伝達細胞は近隣の細胞とともにシグナルに反応することがある(図9.2)。
オートクラインシグナルは、細胞が正しい組織に発達するように、生物の初期発生時にしばしば発生する。 また、オートクラインシグナルは、痛みの感覚や炎症反応を制御している。 さらに、細胞がウイルスに感染した場合、細胞はプログラム細胞死を起こすように自らに信号を送り、その過程でウイルスを殺すことができる。
内分泌シグナル
離れた細胞からのシグナルを内分泌シグナルと呼び、内分泌細胞から発生する。 (体内では、内分泌細胞の多くは甲状腺、視床下部、下垂体などの内分泌腺に存在する)。 この種のシグナルは通常、反応は鈍いものの、効果は長く持続します。 内分泌シグナル伝達で放出されるリガンドはホルモンと呼ばれ、体の一部で作られるが、少し離れた他の体の領域に影響を与えるシグナル伝達分子である(図9.2)
ホルモンは、内分泌細胞とその標的細胞間の大きな距離を血流を介して移動するが、これは体全体に移動するには比較的遅い方法である。 その輸送形態のため、ホルモンは希釈され、標的細胞に作用するときには低濃度で存在することになる。
ダイレクトシグナリング
動物のギャップジャンクションや植物のプラスモデスマータは、隣接する細胞の細胞膜の間の接続部である。 これらの水で満たされたチャンネルは、小さなシグナル分子が2つの細胞間を拡散することを可能にする。 カルシウムイオン(Ca2+)のような小さな分子は細胞間を移動できるが、タンパク質やDNAのような大きな分子はチャネルを通過することができない。 チャネルの特異性により、細胞は独立したまま、迅速かつ容易にシグナルを伝達することができるのです。 直接シグナル伝達を行うことで、ある細胞だけが受け取ったシグナルに対して、複数の細胞が協調して反応することができる。 植物では、プラスモデスマタがいたるところにあり、植物全体を巨大なコミュニケーションネットワークにしている
9.1.2 受容体の種類
受容体は、標的細胞内またはその表面のタンパク質分子で、リガンドと結合する。
内部受容体
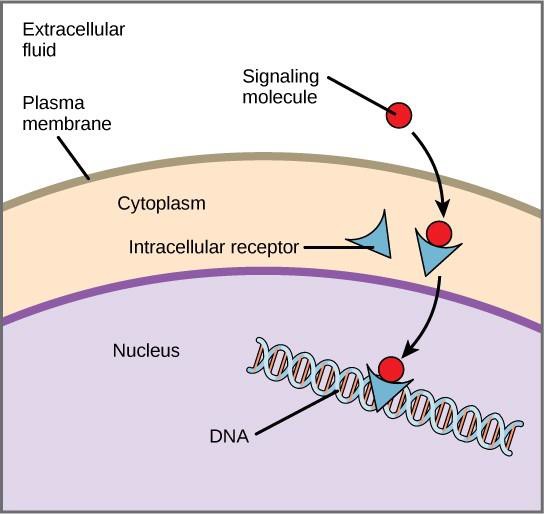
内部受容体は細胞内または細胞質受容体としても知られ、標的細胞の細胞質内に存在し、細胞膜を通過できる疎水性リガンド分子に反応します。
遺伝子発現とは、細胞のDNAにある情報をアミノ酸の配列に変換し、最終的にタンパク質を形成する細胞内のプロセスです。 リガンドが内部の受容体に結合すると、構造変化が起こり、受容体タンパク質上のDNA結合部位が露出します。 リガンドと受容体の複合体は核内に移動し、染色体DNAの特定の制御領域に結合して、転写の開始を促進する(図9.4)。 転写とは、細胞のDNAにある情報を、メッセンジャーRNA(mRNA)という特殊な形態のRNAにコピーするプロセスです。細胞はmRNAの情報を利用して、特定のアミノ酸を正しい順序でつなぎ合わせ、タンパク質を生産します。 このように、リガンドが体内受容体に結合すると、標的細胞の遺伝子発現に直接影響を与えることができます。
細胞表面受容体
細胞表面受容体は膜貫通型受容体としても知られ、外部のシグナル伝達分子と結合する必須タンパク質である。 これらの受容体は細胞膜にまたがっており、細胞外シグナルを細胞間シグナルに変換するシグナル伝達を行う。 (図9.5)。 細胞表面の受容体タンパク質は、細胞が正常に機能するための基本的なものであるため、これらのタンパク質のいずれか1つでも誤作動を起こせば、深刻な結果を招くことは驚くにはあたらないだろう。 9384>
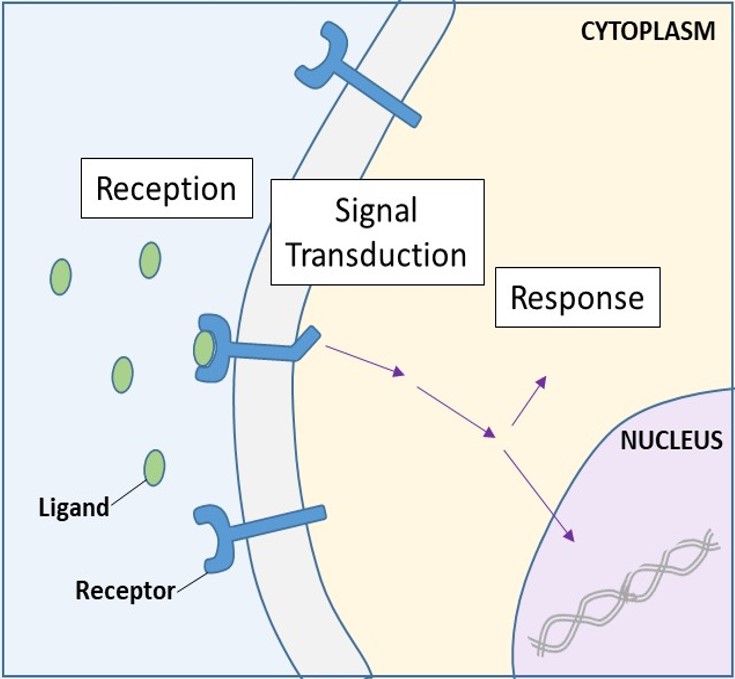
各細胞表面受容体は、外部リガンド結合ドメイン、または細胞外ドメイン、疎水性膜スパン領域、および細胞内ドメインという3つの主要な構成要素を持っています。 細胞表面受容体は多細胞生物におけるシグナル伝達のほとんどに関与している。 酵素結合型受容体、イオンチャンネル結合型受容体、Gタンパク質結合型受容体の3つの一般的なカテゴリーがあります。
酵素結合型受容体は、酵素と結合した細胞内ドメインを持つ細胞表面受容体です。 いくつかのケースでは、受容体の細胞内ドメイン自体が酵素である。 他の酵素結合型受容体は、酵素と直接相互作用する小さな細胞内ドメインを持っています。 酵素結合型受容体は通常、大きな細胞外ドメインと細胞内ドメインを持っているが、膜貫通領域はペプチド鎖の中の1本のαヘリックスからなる。
リガンドが酵素結合型受容体の細胞外ドメインに結合すると、信号が膜を通して伝わり、酵素が活性化される。 酵素の活性化は、最終的に反応を引き起こす細胞内のイベントの連鎖を引き起こす。 細胞外ドメインにシグナル伝達分子が結合すると、受容体は二量体化する。 そして、細胞内ドメインのチロシン残基が自己リン酸化され、下流の細胞応答を誘発する。
酵素結合型受容体の一例として、チロシンキナーゼ受容体がある(図9.6)。 キナーゼとは、ATPから別のタンパク質にリン酸基を転移させる酵素のことである。 チロシンキナーゼ受容体は、リン酸基をチロシン分子に転移させる。 まず、シグナル伝達分子は近くにある2つのチロシンキナーゼ受容体の細胞外ドメインに結合する。 次に2つの隣接する受容体は結合し、あるいは二量化する。 次に、リン酸基が受容体の細胞内ドメイン上のチロシン残基に付加されます(リン酸化)。
上皮成長因子受容体は、このようなシグナル伝達の様式をとる受容体チロシンキナーゼの一例である。 このファミリーのErbBシグナル伝達の欠陥は、多発性硬化症やアルツハイマー病などの神経筋疾患につながる。
イオンチャネル結合型受容体はリガンドに結合して、膜を通して特定のイオンが通過できるようなチャネルを開設する。 このタイプの細胞表面受容体は、疎水性アミノ酸からなる広大な膜貫通領域を持っています。 逆に、チャネルの内側に並ぶアミノ酸は、イオンの通過を可能にするために親水性である。 リガンドがチャネルの細胞外領域に結合すると、タンパク質の構造に構造変化が起こり、ナトリウム、カルシウム、マグネシウム、または水素などのイオンが通過できるようになります(図9.7)。 イオンチャネル結合型受容体の例は神経細胞で見つかります。 神経伝達物質がこれらの受容体に結合すると、構造変化によってナトリウムイオンが細胞膜を横切って流れ、膜電位の変化を引き起こします。
Gタンパク質結合型受容体はリガンドに結合し、関連するGタンパク質を活性化させます。 活性化されたGタンパク質は、次にイオンチャンネルや酵素である近くの膜タンパク質と相互作用する(図9.8)。 すべてのGタンパク質結合型受容体は7つの膜貫通ドメインを持っているが、それぞれの受容体は特定の細胞外ドメインとGタンパク質結合部位を持っている。
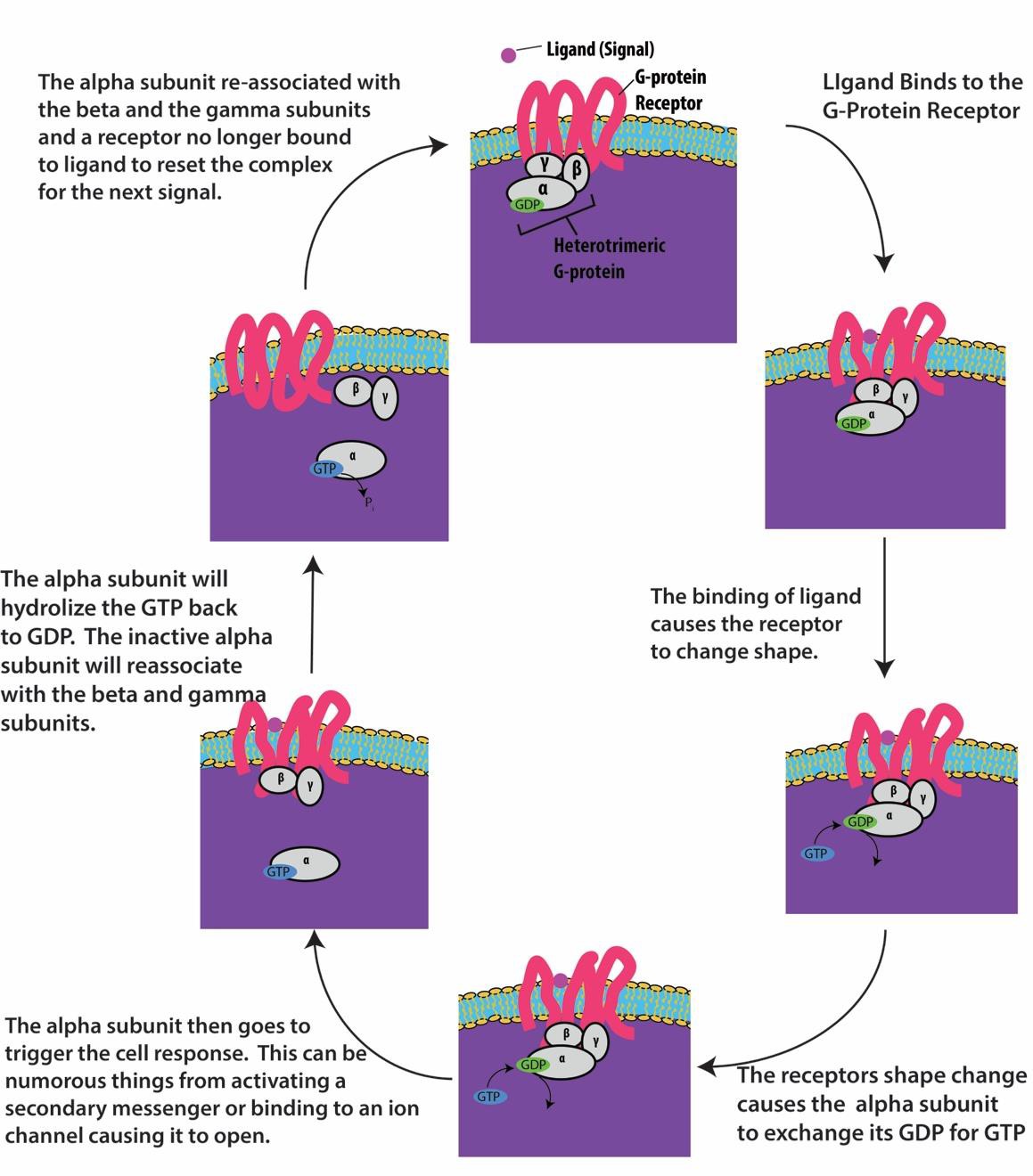
Gタンパク質と結合した受容体を用いた細胞シグナル伝達はサイクルとして起こります。 リガンドが受容体に結合すると、その結果生じる形状の変化によりGタンパク質が活性化され、GDPを放出してGTPを拾います。 その後、Gタンパク質のサブユニットは、αサブユニットとβγサブユニットに分かれる。 これらのGタンパク質の断片の一方または両方が、細胞内の他のタンパク質を活性化することができる。 しばらくすると、Gタンパク質の活性化されたαサブユニット上のGTPはGDPに加水分解され、βγサブユニットは非活性化される。
Gタンパク質と結合した受容体は、視覚伝達、味覚、免疫系や炎症の調節など、多くの生理的プロセスで使用されている。

HER2 は、受容体チロシンキナーゼのひとつです。 ヒト乳がんの30%では、HER2が恒常的に活性化され、細胞分裂が無秩序に行われる。 乳癌の治療に用いられる薬剤であるラパチニブは、受容体が自身をリン酸化するプロセスを阻害し、腫瘍の成長を50%抑制する。 ラパチニブによって阻害されるのは、次のどの段階でしょうか。
- シグナル分子の結合、二量化、および下流の細胞応答
- 二量化、および下流の細胞応答
- ホスファターゼ活性、二量化、および下流の細胞応答
9.1.3 シグナル分子
シグナル伝達細胞によって産生されるリガンドは、標的細胞へ移動して反応を引き起こす化学的シグナルである。 リガンドとして機能する分子の種類は驚くほど多様で、小さなタンパク質から小さなイオンまで多岐にわたる。
Small Hydrophobic Ligands
Small Hydrophobic Ligandsは脂溶性リガンドとも呼ばれ、細胞膜を通して直接拡散し、内部の受容体と相互作用することが可能である。 このクラスのリガンドで重要なのはステロイドホルモンである。 ステロイドは、4つの縮合環を持つ炭化水素骨格を持つ脂質であり、異なるステロイドは、炭素骨格に異なる官能基が結合している。 ステロイドホルモンには、エストロゲンの一種である女性ホルモン、エストラジオール、男性ホルモンであるテストステロン、生体膜の重要な構造成分でステロイドホルモンの前駆体であるコレステロールなどがある(図9.10)。 血液に溶けるためには、疎水性のリガンドが血流に乗って運ばれる間にキャリアータンパク質と結合する必要がある。
水溶性リガンド
水溶性リガンドは極性を持つため、そのままでは細胞膜を通過することができません。 時には大きすぎて、膜をまったく通れないこともある。 その代わりに、ほとんどの水溶性リガンドは細胞表面受容体の細胞外ドメインに結合します(図9.5参照)。 このリガンドグループは非常に多様で、低分子、ペプチド、タンパク質が含まれます。
9.2 | シグナルの伝播
このセクションの終わりまでに、以下のことができるようになります:
- リガンドの結合が細胞全体のシグナル伝達を開始する方法を説明することができる。
- 細胞内シグナルは、リン酸化またはセカンドメッセンジャーの役割によって伝達されることを認識する。
水溶性リガンドがその受容体に結合すると、シグナルは膜を通って細胞質へ伝達される。 このようにシグナルが継続されることをシグナル伝達と呼ぶ(図9.5)。
リガンドがその受容体に結合すると、受容体の細胞内ドメインに影響を与える構造変化が起こります。 これらの構造変化は、細胞内ドメインまたはその関連タンパク質の活性化につながります。 場合によっては、リガンドの結合によって受容体の二量体化が起こり、2つの受容体が互いに結合して二量体と呼ばれる安定した複合体を形成します。 二量体とは、2つの分子(多くの場合同一)が結合して形成される化学化合物のことである。 このように受容体が結合することにより、その細胞内ドメインが密接に接触し、互いに活性化することが可能となる。
9.2.1 シグナル伝達経路とシグナル増幅
シグナル伝達分子はしばしば非常に低濃度で見出されるが、深い効果を生み出すことがある。 リガンドが細胞表面の受容体に結合した後、受容体の細胞内成分の活性化により、シグナル伝達経路またはシグナル伝達カスケードと呼ばれる一連の事象が引き起こされる。 シグナル伝達経路では、セカンドメッセンジャー、酵素、活性化タンパク質が、他のタンパク質やメッセンジャーを活性化する(図9.11)。 シグナル伝達経路の各メンバーは、シグナル増幅と呼ばれるプロセスによって、次のメンバー数千人を活性化することができる。
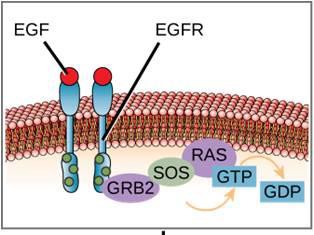
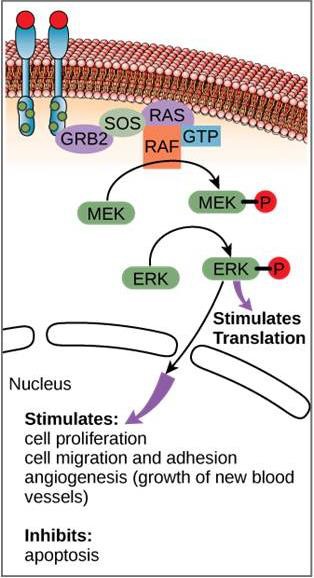
シグナル伝達経路の一例を図9.11に示した。 上皮成長因子(EGF)は、細胞成長、創傷治癒、組織修復の制御に関与するシグナル伝達分子である。 EGFの受容体(EGFR)はチロシンキナーゼである。 活性化されたキナーゼは多くの下流分子をリン酸化し、活性化する。 EGFがEGFRに結合すると、下流のリン酸化イベントのカスケードが細胞の増殖と分裂を促すシグナルを送る。 EGFRが不適切なタイミングで活性化されると、制御不能な細胞増殖(癌)が発生する可能性があります。
特定の癌では、RAS Gタンパク質のGTPase活性が阻害されます。 これは、RASタンパク質がもはやGTPをGDPに加水分解することができないことを意味します。 このことは、下流の細胞イベントにどのような影響を及ぼすでしょうか。
9.2.2 細胞内シグナリングの方法
シグナル伝達経路の誘導は、ある酵素による細胞成分の修飾に依存しています。 経路の次の構成要素を活性化するために起こりうる酵素による修飾は数多くある。 以下は、細胞内シグナル伝達におけるより一般的な事象の一部である。
リン酸化
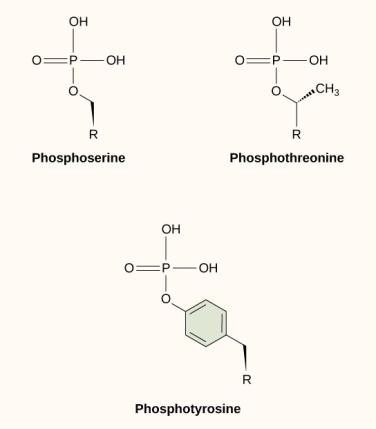
シグナル伝達経路で起こる最も一般的な化学修飾の1つは、リン酸化と呼ばれるプロセスで分子にリン酸基が付加されることである。 リン酸は、GDPまたはGTPを形成するために、GMPのようなヌクレオチドに付加されることができる。 また、タンパク質のセリン、スレオニン、チロシン残基にもしばしばリン酸が付加され、アミノ酸の水酸基と置換される(図9.12)。 リン酸塩の転移は、キナーゼと呼ばれる酵素によって触媒される。 リン酸化は酵素を活性化したり不活性化したりすることがあり、リン酸化の逆である脱リン酸化はその効果を逆転させる。
Second Messenger
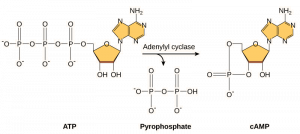
セカンドメッセンジャーとは、シグナル伝達物質が受容体に結合してシグナル伝達が開始された後、それを伝達する小分子を指します。 これらの分子は、特定の細胞タンパク質の挙動を変化させることにより、細胞質を通じて信号を広めるのに役立ちます。 多くの異なる種類の細胞が利用するセカンドメッセンジャーはサイクリック AMP(cAMP)です。 サイクリックAMPは、アデニル酸シクラーゼという酵素によってATPから合成される(図9.13)。 細胞内におけるcAMPの主な役割は、cAMP依存性キナーゼ(Aキナーゼ)と呼ばれる酵素に結合して活性化することである。 A-キナーゼは多くの重要な代謝経路を制御している。 Aキナーゼは、標的タンパク質のセリンおよびスレオニン残基をリン酸化し、その過程でそれらを活性化する。 Aキナーゼは多くの種類の細胞に存在し、それぞれの細胞で標的タンパク質は異なっている。 もう一つのセカンドメッセンジャーはCa2+で、これが放出されて細胞内に溢れることがある。
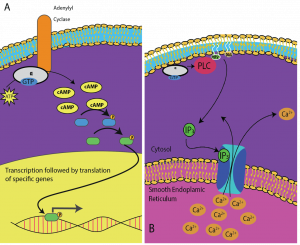
細胞によってcAMPに対する反応は異なる。 図9.14では、Gタンパク質受容体のαサブユニットが2つの異なるタイプのシグナル伝達を活性化している様子が示されている。 最初の図では、αサブユニットによって活性化されると、アデニル酸シクラーゼという酵素からcAMPが生成されます。cAMPは次に、遺伝子の転写に影響を与える他のタンパク質を活性化させます。 2番目の図では、Gタンパク質のαサブユニットがカスケードを引き起こし、平滑小胞体からCa2+が放出される。 この場合、Ca2+は細胞応答を引き起こす二次メッセンジャーである。
9.3 | シグナルへの応答
このセクションの終わりまでに、あなたは次のことができるようになります:
- シグナル伝達経路がタンパク質発現、細胞代謝、および細胞増殖を指示するという事実を認識する。
- 健康な生物の発生と維持におけるアポトーシスの役割を認識する。
シグナル伝達経路を使用して、細胞膜の受容体は細胞に様々な効果をもたらす。 細胞内では、リガンドがその内部の受容体に結合し、細胞のDNAやタンパク質生産機構に直接影響を与えることができます。 シグナル伝達経路の結果は非常に多様で、関与する細胞の種類や外的・内的条件によって異なる。 応答のごく一部を以下に説明する。
9.3.1 シグナル伝達経路に対する応答
Gene Expression
いくつかのシグナル伝達経路は、RNAの転写を調節する。
細胞代謝の増加
アドレナリンによって筋肉細胞のβアドレナリン受容体が活性化されると、細胞内のサイクリックAMPが増加する。 アドレナリンは副腎から分泌されるホルモンで、短期的な緊急事態に体を備える。
細胞成長
細胞シグナル伝達経路も、細胞分裂に大きな役割を担っている。 細胞は、他の細胞からのシグナルによって刺激されない限り、通常は分裂しない。 細胞の成長を促進するリガンドは成長因子と呼ばれる。
細胞死
細胞が損傷したり、不要になったり、生物にとって潜在的に危険な状態になると、細胞はプログラム細胞死、すなわちアポトーシスを引き起こす機構を起動させることができる。 アポトーシスにより、細胞は、細胞内部から潜在的に有害な分子が放出されるのを防ぐ制御された方法で死滅することができる。 しかし、ウイルス感染や癌による無秩序な細胞分裂などの場合、細胞の正常なチェック・アンド・バランスが機能しなくなることがある。 また、外部からのシグナル伝達によってもアポトーシスが引き起こされることがある。 例えば、ほとんどの正常な動物細胞は、動物細胞を構造的に支える糖タンパク質のネットワークである細胞外マトリックスと相互作用する受容体を持っている。 細胞外マトリックスに細胞内受容体が結合すると、細胞内でシグナル伝達カスケードが開始される。 しかし、細胞が細胞外マトリックスから離れると、シグナル伝達は停止し、細胞はアポトーシスを起こす。
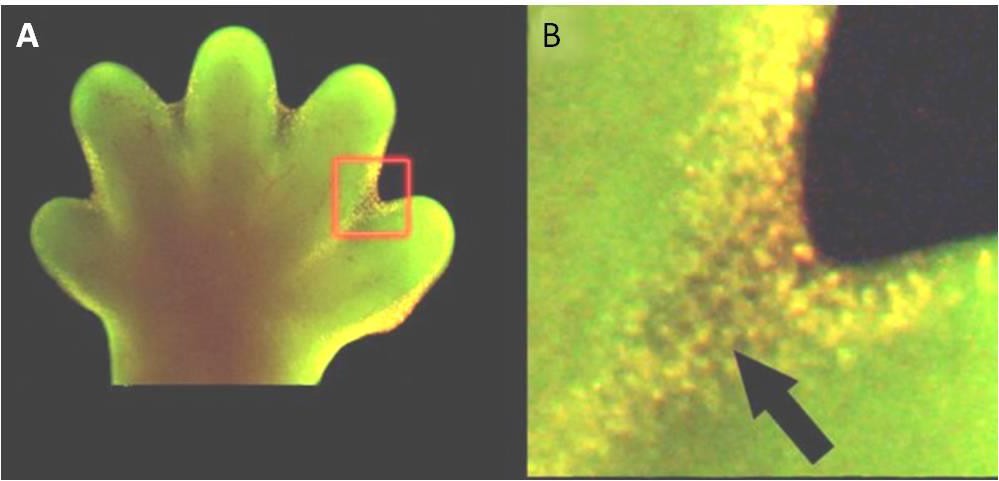
アポトーシスは、通常の胚発生にも不可欠である。 例えば脊椎動物では、発生の初期段階において、個々の指と指の間に網目状の組織が形成される(図9.15)。 正常な発生過程において、これらの不要な細胞は除去され、完全に分離した指や足指が形成されるようになる必要がある。
9.3.2 シグナル伝達経路の終了
腫瘍細胞でしばしば見られる異常なシグナル伝達は、適切なタイミングでのシグナル終了がシグナルの開始と同じくらい重要であることを証明するものである。 特定のシグナルを止める方法のひとつは、リガンドを分解したり、リガンドが受容体にアクセスできなくなるように除去したりすることです。 エストロゲンやテストステロンなどの疎水性ホルモンが、長期間にわたる事象を引き起こす理由の1つは、キャリアタンパク質と結合するためである。
細胞内では、多くの異なる酵素が、シグナル伝達カスケードから生じる細胞の修飾を逆転させる。 例えば、ホスファターゼは、脱リン酸化と呼ばれるプロセスで、キナーゼによってタンパク質に結合したリン酸基を取り除く酵素である。cAMPはホスホジエステラーゼによってAMPに分解され、それによってそのシグナルが停止する。 同様に、貯蔵されたカルシウムの放出は、細胞膜に存在するCa2+ポンプによって逆転される
。