
Digite seus exemplos aqui.
- 9.1 Moléculas de Sinalização e Receptores Celulares
- 9.2 Propagação do Sinal
- 9.3 Resposta ao Sinal
Imagine como seria a vida se você e as pessoas ao seu redor não pudessem se comunicar. Você não seria capaz de expressar seus desejos aos outros, nem poderia fazer perguntas para saber mais sobre seu ambiente. A organização social depende da comunicação entre os indivíduos que compõem essa sociedade.
Como com as pessoas, é vital que as células individuais sejam capazes de interagir com seu ambiente e umas com as outras. Isto é verdade quer uma célula esteja crescendo sozinha num tanque ou seja uma das muitas células que formam um organismo maior. Para responder adequadamente aos estímulos externos, as células desenvolveram mecanismos complexos de comunicação para que possam receber uma mensagem, transferir a informação através da membrana plasmática e depois produzir mudanças dentro da célula em resposta à mensagem.
Em organismos multicelulares, as células enviam e recebem constantemente mensagens químicas para coordenar as ações de outros órgãos, tecidos e células. A capacidade de enviar mensagens de forma rápida e eficiente permite que as células coordenem e ajustem as suas funções. Enquanto a necessidade de comunicação celular em organismos maiores parece óbvia, mesmo organismos unicelulares comunicam entre si. As células de levedura sinalizam umas às outras para ajudar no acasalamento. Algumas formas de bactérias coordenam suas ações a fim de formar grandes complexos chamados biofilmes ou para organizar a produção de toxinas para remover organismos concorrentes. A capacidade das células se comunicarem através de sinais químicos originados em células únicas e foi essencial para a evolução de organismos multicelulares. A comunicação eficiente e sem erros é vital para toda a vida.
- 9.1 | Sinalizando Moléculas e Receptores Celulares
- 9.1.1 Formas de Sinalização
- Sinalização parácrina
- Autocrina de sinalização
- Sinalização endócrina
- Sinalização Direta
- 9.1.2 Tipos de Receptores
- Receptores internos
- Receptores de superfície celular
- 9.1.3 Moléculas de Sinalização
- Ligantes hidrofóbicos pequenos
- Ligantes solúveis em água
- 9.2 | Propagação do Sinal
- 9.2.1 Caminhos de Sinalização e Amplificação de Sinal
- 9.2.2 Métodos de Sinalização Intracelular
- Fosforilação
- Segundo Mensageiros
- 9.3 |Resposta ao Sinal
- 9.3.1 Respostas à via de sinalização
- Expressão do gene
- Increase in Cellular Metabolism
- Celular Growth
- Morte celular
- 9.3.2 Término das vias de sinalização
9.1 | Sinalizando Moléculas e Receptores Celulares
Ao final desta seção, você será capaz de:
- Descrever quatro tipos de sinalização encontrados em organismos multicelulares.
- Comparar receptores internos com receptores de superfície celular.
- Reconhecer a relação entre a química de um ligando e seu mecanismo de ação.
Existem dois tipos de comunicação no mundo das células vivas. A comunicação entre células é chamada de sinalização intercelular, e a comunicação dentro de uma célula é chamada de sinalização intracelular. Uma maneira fácil de lembrar a distinção é entendendo que o prefixo entre significa “entre” (uma auto-estrada interestadual cruza entre estados) e intra- significa “dentro” (um IV significa intravenoso ou “dentro da veia”).
Sinais químicos são liberados por uma célula de sinalização e recebidos por uma célula alvo. As células alvo têm proteínas chamadas receptores, que se ligam às moléculas de sinalização e causam uma resposta. As moléculas sinalizadoras que se ligam aos receptores são chamadas de ligandos. Os ligandos e receptores são específicos um do outro; um receptor normalmente liga-se apenas ao seu ligando específico. Entretanto, existem diferentes tipos de sinalização.
9.1.1 Formas de Sinalização
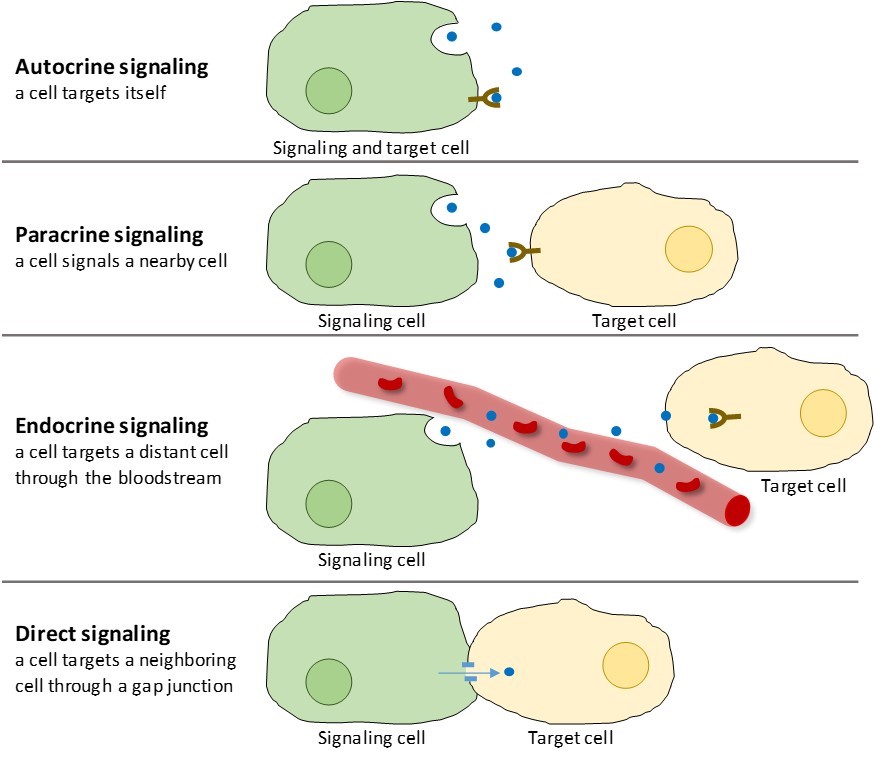
Existem quatro categorias de sinalização química encontradas em organismos multicelulares: sinalização autocrina, sinalização parácrina, sinalização endócrina, e sinalização direta através de junções de fendas (Figura 9.2). A principal diferença entre as diferentes categorias de sinalização é a distância que o sinal viaja para alcançar a célula alvo.
Sinalização parácrina
Sinais que atuam localmente entre células que estão próximas umas das outras são chamados de sinais parácrinos. Os sinais parácrinos movem-se por difusão através da matriz extracelular (Figura 9.2). Estes tipos de sinais normalmente provocam respostas rápidas que duram apenas um curto período de tempo. A fim de manter a resposta localizada, os ligandos parácrinos são geralmente rapidamente degradados pelas enzimas ou removidos pelas células vizinhas. A remoção dos sinais restabelece o gradiente de concentração da molécula do sinal, permitindo que eles se difundam rapidamente através do espaço intracelular se liberados novamente.
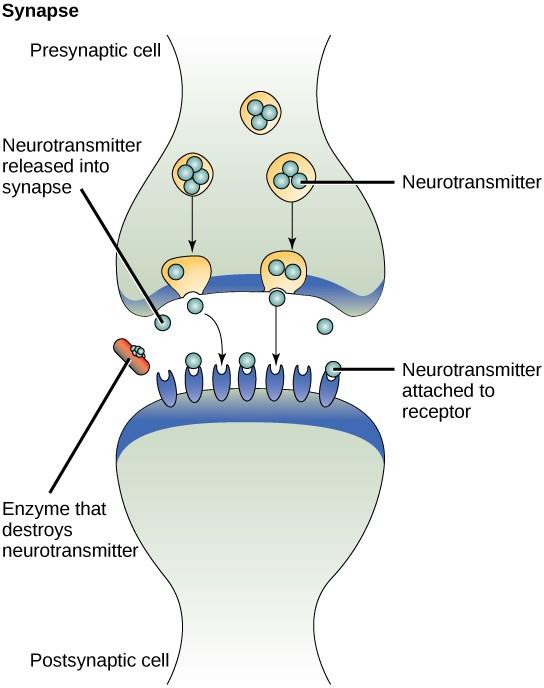
Um exemplo de sinalização parácrina é a transferência de sinais entre células nervosas. O pequeno espaço entre as células nervosas onde ocorre a transmissão do sinal é chamado de sinapse. Os sinais são propagados ao longo das células nervosas através de impulsos elétricos de movimento rápido. Quando esses impulsos chegam ao final de uma célula nervosa, ligandos químicos chamados neurotransmissores são liberados na sinapse pela célula pré-sináptica (a célula que emite o sinal). Os neurotransmissores difundem-se através da sinapse (Figura 9.3). A pequena distância entre as células nervosas permite que o sinal viaje rapidamente, o que permite uma resposta imediata, como, por exemplo, “tire a mão do fogão”! Quando o neurotransmissor liga o receptor na superfície da célula pós-sináptica, o próximo impulso elétrico é lançado. Os neurotransmissores são degradados rapidamente ou são reabsorvidos pela célula pré-sináptica para que a célula nervosa receptora possa recuperar rapidamente e estar preparada para responder rapidamente ao próximo sinal sináptico.
Autocrina de sinalização
Quando uma célula responde à sua própria molécula de sinalização, ela é chamada de autocrina de sinalização (auto = “self”). A sinalização autocrítica ocorre frequentemente com outros tipos de sinalização. Por exemplo, quando um sinal parácrino é liberado, a célula de sinalização pode responder ao sinal junto com seus vizinhos (Figura 9.2).
Autocrina de sinalização freqüentemente ocorre durante o desenvolvimento precoce de um organismo para assegurar que as células se desenvolvam nos tecidos corretos. A sinalização autócrina também regula a sensação de dor e as respostas inflamatórias. Além disso, se uma célula está infectada com um vírus, a célula pode sinalizar a si mesma para sofrer a morte programada da célula, matando o vírus no processo.
Sinalização endócrina
Sinais de células distantes são chamados de sinais endócrinos, e se originam de células endócrinas. (No corpo, muitas células endócrinas estão localizadas em glândulas endócrinas, como a glândula tiróide, o hipotálamo e a glândula pituitária). Estes tipos de sinais geralmente produzem uma resposta mais lenta, mas têm um efeito mais duradouro. Os ligandos liberados na sinalização endócrina são chamados hormônios, sinalizando moléculas que são produzidas em uma parte do corpo, mas afetam outras regiões do corpo a alguma distância (Figura 9.2).
Hormônios percorrem as grandes distâncias entre as células endócrinas e suas células-alvo através da corrente sanguínea, o que é uma forma relativamente lenta de se mover por todo o corpo. Devido à sua forma de transporte, as hormonas são diluídas e estão presentes em baixas concentrações quando actuam sobre as células alvo. Isto é diferente da sinalização parácrina, na qual as concentrações locais de moléculas de sinalização podem ser muito altas.
Sinalização Direta
Junções de gotas em animais e plasmodesmados em plantas são conexões entre as membranas plasmáticas de células vizinhas. Estes canais cheios de água permitem que pequenas moléculas de sinalização se difundam entre as duas células. Pequenas moléculas, tais como íons de cálcio (Ca2+), são capazes de se mover entre as células, mas moléculas grandes como proteínas e DNA não cabem através dos canais. A especificidade dos canais assegura que as células permanecem independentes, mas podem transmitir sinais rápida e facilmente. A sinalização directa permite a um grupo de células coordenar a sua resposta a um sinal que apenas uma delas pode ter recebido. Nas plantas, os plasmodesmados são omnipresentes, transformando toda a planta numa gigantesca rede de comunicação.
9.1.2 Tipos de Receptores
Receptores são moléculas proteicas na célula alvo ou na sua superfície que se ligam aos ligandos. Existem dois tipos de receptores, receptores internos e receptores de superfície celular.
Receptores internos
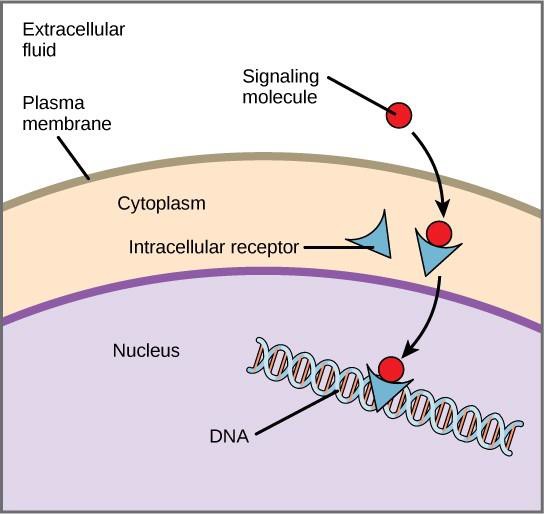
Receptores internos, também conhecidos como receptores intracelulares ou citoplasmáticos, são encontrados no citoplasma das células alvo e respondem a moléculas de ligantes hidrofóbicos que são capazes de viajar através da membrana plasmática. Uma vez dentro da célula, muitas dessas moléculas se ligam a proteínas que atuam como reguladores da síntese do mRNA (transcrição) para mediar a expressão gênica.
Expressão do gene é o processo celular de transformar a informação no DNA de uma célula em uma seqüência de aminoácidos, que finalmente forma uma proteína. Quando o ligante se liga ao receptor interno, uma mudança conformacional é desencadeada que expõe um local de ligação de DNA na proteína receptora. O complexo ligante-receptor move-se para o núcleo, depois liga-se a regiões reguladoras específicas do DNA cromossômico e promove o início da transcrição (Figura 9.4). Transcrição é o processo de copiar a informação no DNA de uma célula em uma forma especial de RNA chamado RNA mensageiro (mRNA); a célula usa a informação no mRNA para ligar aminoácidos específicos na ordem correta, produzindo uma proteína. Assim, quando um ligante se liga a um receptor interno, ele pode influenciar diretamente a expressão gênica na célula alvo.
Receptores de superfície celular
Receptores de superfície celular, também conhecidos como receptores transmembrana, são proteínas integrais que se ligam a moléculas de sinalização externa. Estes receptores atravessam a membrana plasmática e realizam a transdução de sinal, na qual um sinal extracelular é convertido em um sinal intercelular. (Figura 9.5). Como as proteínas receptoras da superfície celular são fundamentais para o funcionamento normal das células, não deve ser surpresa que um mau funcionamento em qualquer uma destas proteínas possa ter consequências graves. Erros nas estruturas proteicas de certas moléculas receptoras têm demonstrado um papel na hipertensão (pressão alta), asma, doença cardíaca e câncer.
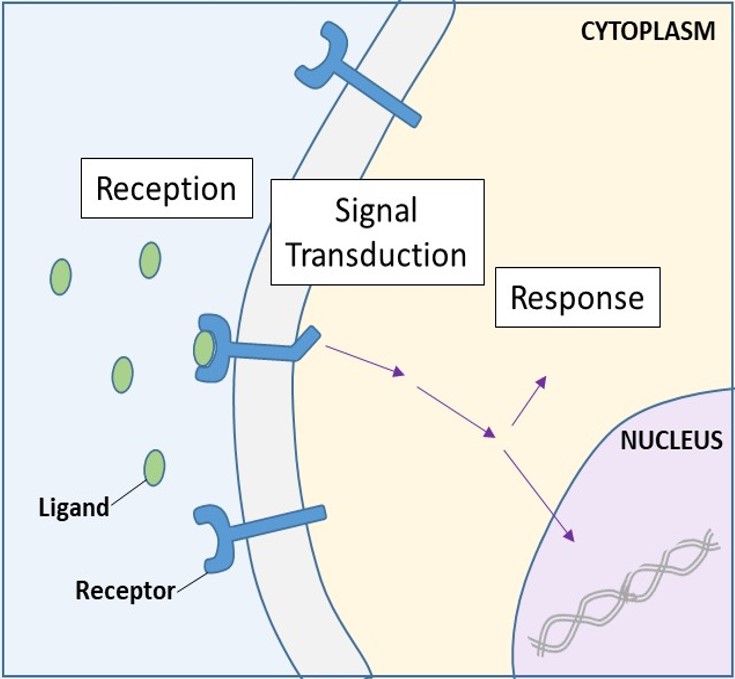 Figure 9.5 Moléculas de sinalização hidrofílica normalmente funcionam ligando-se à porção extracelular de uma proteína receptora. O sinal é então transduzido através da membrana.
Figure 9.5 Moléculas de sinalização hidrofílica normalmente funcionam ligando-se à porção extracelular de uma proteína receptora. O sinal é então transduzido através da membrana.Cada receptor de superfície celular tem três componentes principais: um domínio ligante externo, ou domínio extracelular; uma região hidrofóbica de membranas; e um domínio intracelular. Os receptores de superfície celular estão envolvidos na maior parte da sinalização em organismos multicelulares. Há três categorias gerais de receptores de superfície celular: receptores ligados à enzima, receptores ligados ao canal iônico e receptores ligados à proteína G. Os receptores ligados à enzima são receptores de superfície celular com domínios intracelulares que estão associados a uma enzima. Em alguns casos, o domínio intracelular do próprio receptor é uma enzima. Outros receptores ligados a enzimas têm um pequeno domínio intracelular que interage diretamente com uma enzima. Os receptores ligados à enzima normalmente têm grandes domínios extracelulares e intracelulares, mas a região de extensão da membrana consiste de uma única hélice alfa-helix na cadeia de peptídeos.
Quando um ligando se liga ao domínio extracelular de um receptor ligado à enzima, um sinal é transferido através da membrana, ativando a enzima. A ativação da enzima desencadeia uma cadeia de eventos dentro da célula que eventualmente leva a uma resposta.
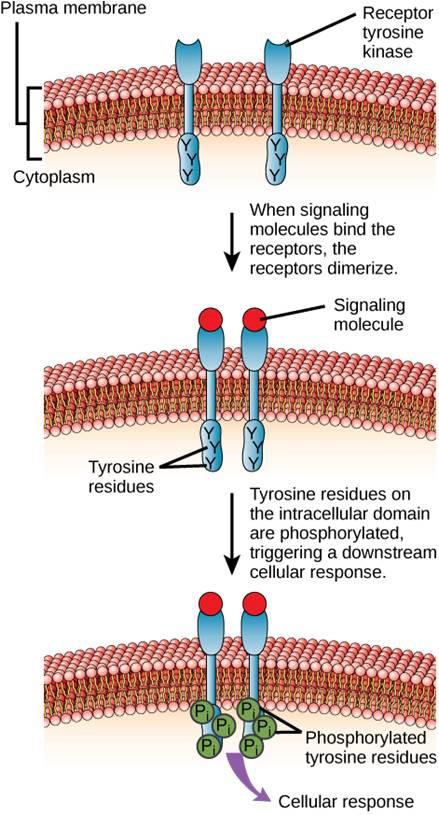
Um exemplo de um receptor ligado a uma enzima é o receptor de tirosina quinase (Figura 9.6). Uma quinase é uma enzima que transfere grupos de fosfato de ATP para outra proteína. O receptor de tirosina quinase transfere grupos de fosfato para moléculas de tirosina. Primeiro, as moléculas sinalizadoras ligam-se ao domínio extracelular de dois receptores de tirosina quinase próximos. Os dois receptores vizinhos então se ligam, ou dimerizam. Os fosfatos são então adicionados aos resíduos de tirosina no domínio intracelular dos receptores (fosforilação). Os resíduos fosforilados podem então transmitir o sinal para o próximo mensageiro dentro do citoplasma.
Receptores de fator de crescimento da epiderme são um exemplo de receptores de tirosina kinases que seguem este modo de sinalização. Defeitos na sinalização ErbB nesta família podem levar a doenças neuromusculares tais como esclerose múltipla e doença de Alzheimer.
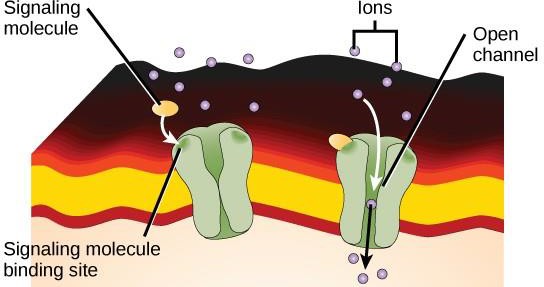
Receptores ligados a um ligando de iões ligam-se a um ligando e abrem um canal através da membrana que permite a passagem de iões específicos. Este tipo de receptor de superfície celular tem uma extensa região de membranas com aminoácidos hidrofóbicos. Por outro lado, os aminoácidos que revestem o interior do canal são hidrofílicos para permitir a passagem de íons. Quando um ligante se liga à região extracelular do canal, há uma mudança conformacional na estrutura da proteína que permite a passagem de íons como sódio, cálcio, magnésio ou hidrogênio (Figura 9.7).
Receptores ligados à proteína G ligam-se a um ligante e ativam uma proteína G associada. A proteína G ativada interage então com uma proteína da membrana próxima, que pode ser um canal iônico ou uma enzima (Figura 9.8). Todos os receptores ligados à proteína G têm sete domínios transmembrana, mas cada receptor tem um domínio extracelular específico e um site de ligação à proteína G.
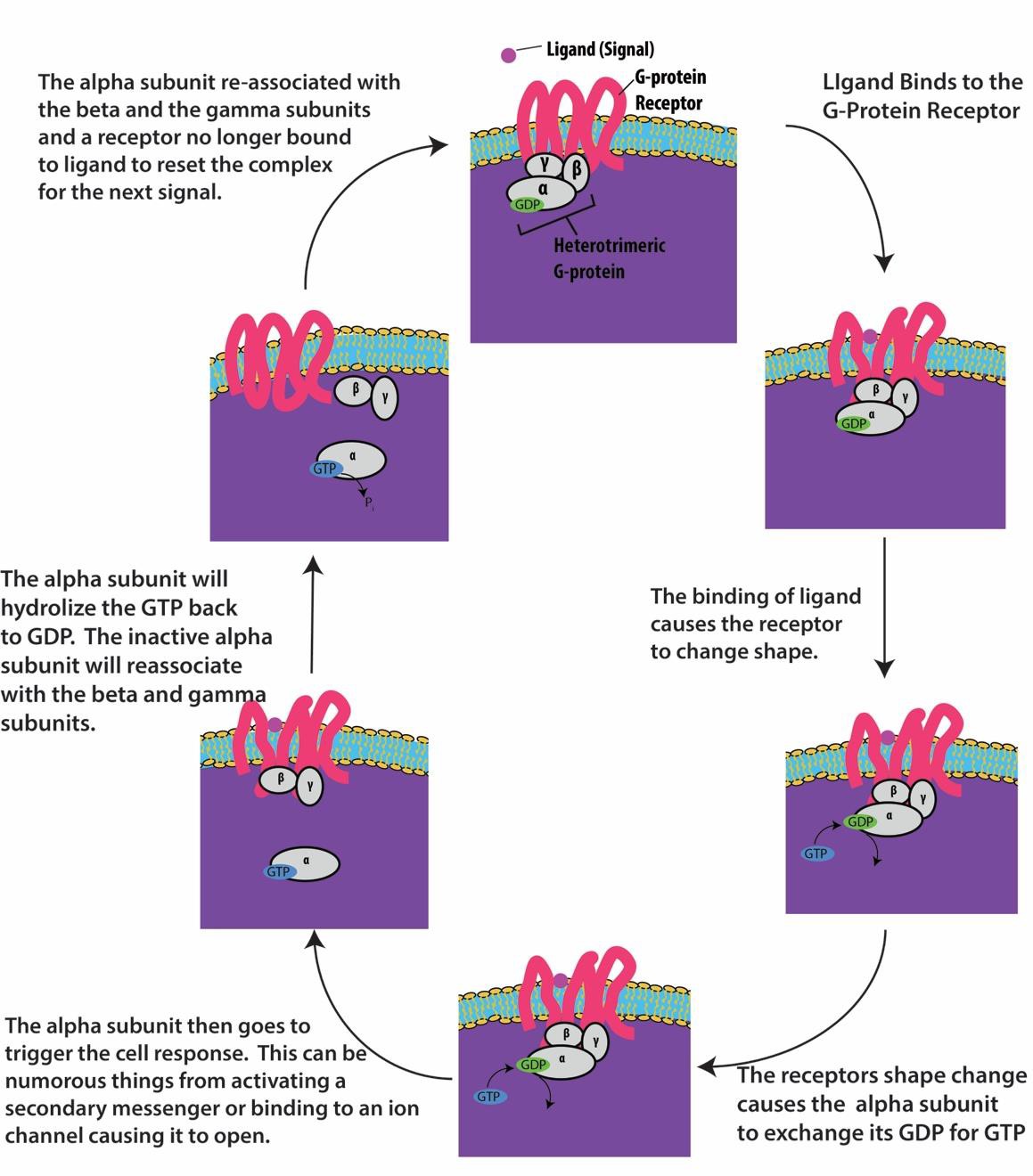
Sinalização celular usando receptores ligados à proteína G ocorre como um ciclo. Uma vez que o ligante se liga ao receptor, a mudança de forma resultante ativa a proteína G, que libera o GDP e pega o GTP. As subunidades da proteína G dividem-se então em α e βγ subunidades. Um ou ambos esses fragmentos de proteína G podem ser capazes de ativar outras proteínas na célula. Após algum tempo, o GTP na subunidade ativa α da proteína G é hidrolisado ao GDP e a subunidade βγ é desativada. As subunidades se associam novamente para formar a proteína G inativa e o ciclo começa novamente (Figura 9.8).
receptores ligados à proteína G são usados em muitos processos fisiológicos, incluindo aqueles para a transdução da visão, sabor e regulação do sistema imunológico e inflamação.

HER2 é uma tirosina kinase receptora. Em 30% dos cancros mamários humanos, HER2 é permanentemente ativado, resultando em divisão celular não regulada. O Lapatinib, uma droga usada para tratar o câncer de mama, inibe o processo pelo qual o próprio receptor fosforilatos, reduzindo assim o crescimento tumoral em 50%. Qual dos seguintes passos seria inibido pelo Lapatinib?
- Atividade da molécula de sinalização, dimerização e a resposta celular a jusante
- Dimerização, e a resposta celular a jusante
- A resposta celular a jusante
- Atividade da fosfatase, dimerização, e a resposta celular a jusante
9.1.3 Moléculas de Sinalização
Produzidos pelas células de sinalização, os ligandos são sinais químicos que viajam para as células alvo e causam uma resposta. Os tipos de moléculas que servem como ligandos são incrivelmente variados e variam de pequenas proteínas a pequenos iões. Os ligandos são categorizados como pequenos ligandos hidrofóbicos, que podem atravessar membranas plasmáticas, ou ligandos solúveis em água, que não podem.
Ligantes hidrofóbicos pequenos
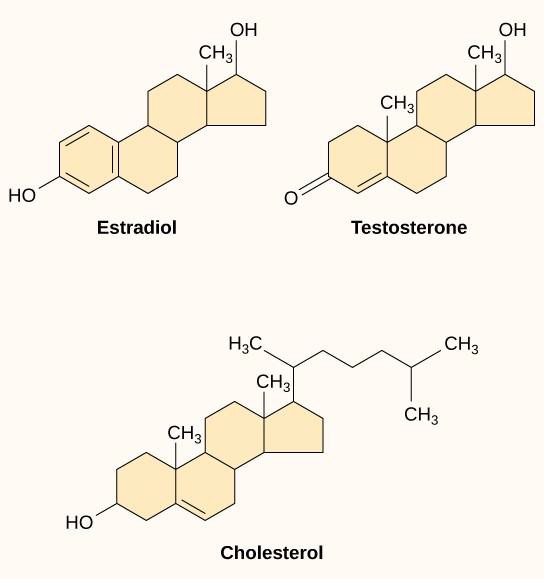
Ligantes hidrofóbicos pequenos, também chamados de ligantes lipossolúveis, podem se difundir diretamente através da membrana plasmática e interagir com os receptores internos. Membros importantes desta classe de ligandos são os hormônios esteróides. Os esteróides são lipídios que têm um esqueleto de hidrocarbonetos com quatro anéis fundidos; esteróides diferentes têm grupos funcionais diferentes ligados ao esqueleto de carbono. Os hormônios esteróides incluem o hormônio sexual feminino estradiol, que é um tipo de estrogênio; o hormônio sexual masculino testosterona; e o colesterol, que é um importante componente estrutural das membranas biológicas e um precursor dos hormônios esteróides (Figura 9.10). Outros hormônios hidrofóbicos incluem hormônios tireoidianos e vitamina D. Para serem solúveis no sangue, os ligantes hidrofóbicos devem se ligar às proteínas transportadoras enquanto são transportados pela corrente sanguínea.
Ligantes solúveis em água
Ligantes solúveis em água são polares, não podem passar através da membrana plasmática sem ajuda. Por vezes são demasiado grandes para passar através da membrana. Em vez disso, a maioria dos ligandos solúveis em água ligam-se ao domínio extracelular dos receptores de superfície celular (ver Figura 9.5). Este grupo de ligandos é bastante diversificado e inclui pequenas moléculas, peptídeos e proteínas.
9.2 | Propagação do Sinal
Ao final desta seção, você será capaz de:
- Explicar como a ligação de um ligando inicia a transdução do sinal através de uma célula.
- Reconsiderar que sinais intracelulares são transmitidos pelo papel da fosforilação ou segundos mensageiros.
Após a ligação de um ligando hidrossolúvel ao seu receptor, o sinal é transmitido através da membrana e para o citoplasma. A continuação de um sinal desta forma é chamada de transdução de sinal (Figura 9.5). A transdução de sinal só ocorre com receptores de superfície celular, pois os receptores internos são capazes de entrar na célula.
Quando um ligando se liga ao seu receptor, ocorrem mudanças conformacionais que afetam o domínio intracelular do receptor. Essas alterações conformacionais levam à ativação do domínio intracelular ou de suas proteínas associadas. Em alguns casos, a ligação do ligante provoca a dimerização do receptor, o que significa que dois receptores se ligam um ao outro para formar um complexo estável chamado dimer. Um dímero é um composto químico formado quando duas moléculas (muitas vezes idênticas) se juntam. A ligação dos receptores desta maneira permite que seus domínios intracelulares entrem em contato próximo e ativem-se mutuamente.
9.2.1 Caminhos de Sinalização e Amplificação de Sinal
As moléculas de sinalização são frequentemente encontradas em concentrações muito baixas, mas podem produzir efeitos profundos. Depois que o ligante se liga ao receptor de superfície celular, a ativação dos componentes intracelulares do receptor desencadeia uma cadeia de eventos que é chamada de caminho de sinalização ou uma cascata de sinalização. Em uma via de sinalização, os segundos mensageiros, enzimas e/ou proteínas ativadas ativam outras proteínas ou mensageiros (Figura 9.11). Cada membro da via pode ativar milhares do próximo membro da via em um processo chamado amplificação de sinal. Como o sinal é amplificado a cada passo, uma resposta muito grande pode ser gerada de um único receptor ligando um ligando.
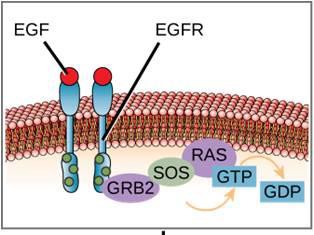
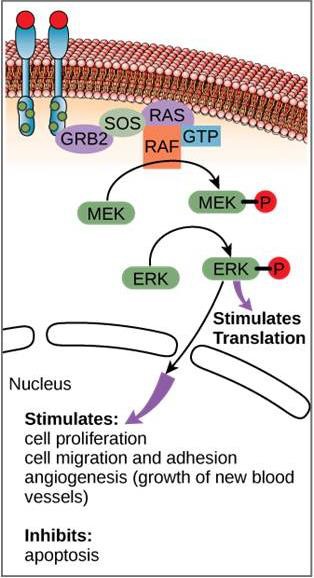
Um exemplo de caminho de sinalização é mostrado na Figura 9.11. O fator de crescimento epidérmico (EGF) é uma molécula de sinalização que está envolvida na regulação do crescimento celular, na cicatrização de feridas e na reparação de tecidos. O receptor do EGF (EGFR) é uma tirosina quinase. Um fosforilatos cinase ativado e ativa muitas moléculas a jusante. Quando o EGF se liga ao EGFR, uma cascata de eventos de fosforilação a jusante sinaliza o crescimento e a divisão da célula. Se o EGFR for ativado em momentos inapropriados, pode ocorrer crescimento celular descontrolado (câncer).
Em determinados cancros, a atividade da proteína GTPase do RAS G é inibida. Isto significa que a proteína RAS não pode mais hidrolisar o GTP no PIB. Que efeito isto teria em eventos celulares a jusante?
9.2.2 Métodos de Sinalização Intracelular
A indução de uma via de sinalização depende da modificação de um componente celular por uma enzima. Há numerosas modificações enzimáticas que podem ocorrer para ativar o próximo componente da via. A seguir estão alguns dos eventos mais comuns na sinalização intracelular.
Fosforilação
Uma das modificações químicas mais comuns que ocorrem nas vias de sinalização é a adição de um grupo de fosfato a uma molécula em um processo chamado fosforilação. O fosfato pode ser adicionado a um nucleotídeo como o GMP para formar o GDP ou GTP. Os fosfatos também são frequentemente adicionados aos resíduos de proteínas serina, treonina e tirosina, onde substituem o grupo hidroxila do aminoácido (Figura 9.12). A transferência do fosfato é catalisada por uma enzima chamada quinase. A fosforilação pode ativar ou inativar enzimas, e a reversão da fosforilação, desfosforilação, reverterá o efeito.
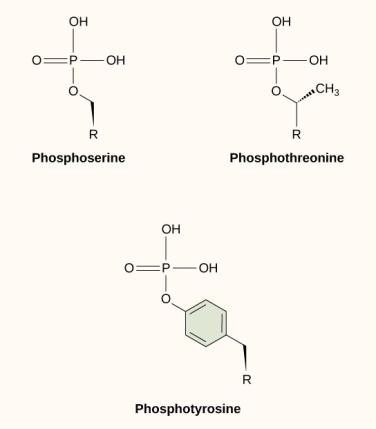 Figure 9.12 Na fosforilação da proteína, um grupo fosfato (PO4-3) é adicionado aos resíduos dos aminoácidos serina, treonina, ou tirosina. O grupo fosfato é adicionado por uma quinase. O ATP é frequentemente utilizado como substrato para adicionar o grupo fosfato a estes aminoácidos. O grupo fosfato frequentemente resulta em uma mudança de forma na proteína que pode ativar ou desligar a função da proteína.
Figure 9.12 Na fosforilação da proteína, um grupo fosfato (PO4-3) é adicionado aos resíduos dos aminoácidos serina, treonina, ou tirosina. O grupo fosfato é adicionado por uma quinase. O ATP é frequentemente utilizado como substrato para adicionar o grupo fosfato a estes aminoácidos. O grupo fosfato frequentemente resulta em uma mudança de forma na proteína que pode ativar ou desligar a função da proteína.Segundo Mensageiros
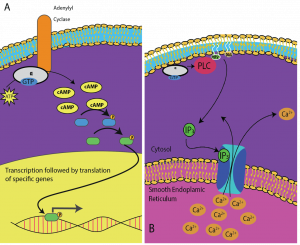
Segundo Mensageiros são pequenas moléculas que propagam um sinal depois de ter sido iniciado pela ligação da molécula sinalizadora ao receptor. Estas moléculas ajudam a propagar um sinal através do citoplasma, alterando o comportamento de certas proteínas celulares. Um segundo mensageiro utilizado por muitos tipos diferentes de células é o AMP cíclico (cAMP). O AMP cíclico é sintetizado pela enzima adeniliclase a partir do ATP (Figura 9.13). O papel principal do AMPciclico nas células é ligar e ativar uma enzima chamada cinase dependente do AMPc (A-kinase). A-quinase regula muitas vias metabólicas vitais: Ela fosforilatos serina e resíduos de treonina das suas proteínas alvo, activando-as no processo. A-quinase é encontrada em muitos tipos diferentes de células, e as proteínas alvo em cada tipo de célula são diferentes. Outro mensageiro secundário é Ca2+, que pode ser liberado para inundar a célula.
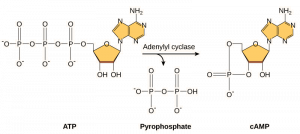
Células diferentes respondem diferentemente ao AMPc. Na Figura 9.14, a subunidade alfa de um receptor de proteína G é mostrada ativando dois tipos diferentes de sinalização. Na primeira imagem, o cAMP é produzido pela enzima adenilato ciclase quando ativado pela subunidade alfa. cAMP então ativa outras proteínas que afetam a transcrição do gene. Na segunda imagem, a subunidade alfa da proteína G dispara uma cascata que libera Ca2+ a partir do retículo endoplasmático liso. Neste caso, Ca2+ é o mensageiro secundário que causa a resposta celular.
9.3 |Resposta ao Sinal
Ao final desta seção, você será capaz de:
- Reconsiderar que as vias de sinalização direcionam a expressão da proteína, o metabolismo celular e o crescimento celular.
- Reconhecer o papel da apoptose no desenvolvimento e manutenção de um organismo saudável.
Usando vias de transdução de sinal, os receptores na membrana plasmática produzem uma variedade de efeitos sobre a célula. Dentro da célula, os ligandos ligam-se aos seus receptores internos, permitindo-lhes afectar directamente o ADN da célula e as máquinas produtoras de proteínas. Os resultados das vias de sinalização são extremamente variados e dependem do tipo de célula envolvida, bem como das condições externas e internas. Uma pequena amostra das respostas é descrita abaixo.
9.3.1 Respostas à via de sinalização
Expressão do gene
Algumas vias de transdução de sinal regulam a transcrição do RNA. Outras regulam a tradução de proteínas.
Increase in Cellular Metabolism
A activação de β-adrenergic receptors in muscle cells by adrenaline leads to an increase in cyclic AMP inside the cell. A adrenalina é um hormônio produzido pela glândula adrenal que prepara o corpo para emergências de curto prazo.
Celular Growth
Caminhos de sinalização celular também desempenham um papel importante na divisão celular. As células normalmente não se dividem a menos que sejam estimuladas por sinais de outras células. Os ligandos que promovem o crescimento celular são chamados fatores de crescimento. A maioria dos fatores de crescimento ligam-se aos receptores de superfície celular que estão ligados à tirosina kinases.
Morte celular
Quando uma célula é danificada, supérflua, ou potencialmente perigosa para um organismo, uma célula pode iniciar um mecanismo para desencadear a morte celular programada, ou apoptose. A apoptose permite que uma célula morra de uma maneira controlada que previne a liberação de moléculas potencialmente prejudiciais do interior da célula. No entanto, em alguns casos, tais como uma infecção viral ou uma divisão celular não controlada devido ao câncer, as verificações e equilíbrios normais da célula falham. A sinalização externa também pode iniciar a apoptose. Por exemplo, a maioria das células animais normais tem receptores que interagem com a matriz extracelular, uma rede de glicoproteínas que fornece suporte estrutural para as células animais. A ligação dos receptores celulares à matriz extracelular inicia uma cascata de sinalização dentro da célula. Entretanto, se a célula se afasta da matriz extracelular, a sinalização cessa, e a célula sofre apoptose. Este sistema ajuda a evitar que as células viajem pelo corpo e proliferem fora de controle, como acontece com células tumorais que se metástase.
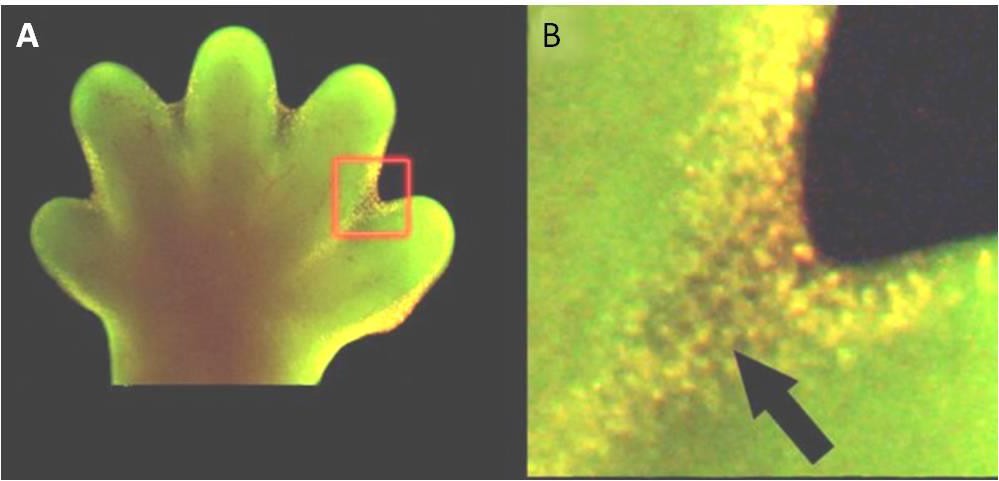
Apoptose também é essencial para o desenvolvimento embriológico normal. Em vertebrados, por exemplo, os estágios iniciais de desenvolvimento incluem a formação de tecido semelhante a uma teia entre dedos individuais e dedos dos pés (Figura 9.15). No decurso do desenvolvimento normal, estas células desnecessárias devem ser eliminadas, permitindo a formação de dedos totalmente separados entre si. Um mecanismo de sinalização celular desencadeia a apoptose, que destrói as células entre os dígitos em desenvolvimento.
9.3.2 Término das vias de sinalização
A aberração da sinalização frequentemente observada nas células tumorais é a prova de que o término de um sinal no momento apropriado pode ser tão importante quanto o início de um sinal. Um método de parar um sinal específico é degradar o ligando ou removê-lo para que ele não possa mais acessar seu receptor. Uma razão para que hormonas hidrofóbicas como o estrogénio e a testosterona desencadeiem eventos duradouros é porque se ligam a proteínas portadoras. Estas proteínas permitem que as moléculas insolúveis sejam solúveis no sangue, mas também protegem os hormônios da degradação pelas enzimas circulantes.
No interior da célula, muitas enzimas diferentes revertem as modificações celulares que resultam das cascatas sinalizadoras. Por exemplo, as fosfatases são enzimas que removem o grupo fosfato ligado às proteínas pelas kinases num processo chamado desfosforização. cAMP é degradado em AMP pela fosfodiesterase, terminando assim o seu sinal. Da mesma forma, a liberação de reservas de cálcio é revertida por bombas de Ca2+ que estão localizadas nas membranas celulares.