Kokeen yksityiskohtainen kuvaus
- Johdanto
- Materiaalit ja menetelmät
- Kysymykset jatkopohdintaa ja keskustelua varten
- Viitteet ja linkit
- Työkaluja opiskelijoiden oppimistulosten arviointiin
- Työkaluja tämän kokeen formatiiviseen arviointiin
Esittely:
Tässä tutkimuksessa tarkastelemme resursseista käytävän kilpailun vaikutuksia lisääntymistulokseen kahden loisampiaislajin sisällä ja niiden välillä.
Laboratorion tavoitteet:
Tämän laboratorion päätteeksi opiskelijat osaavat…
- kuvaamaan Nasonia vitripenniksen ja Melittobia digitatan elinkiertoa,
- selittämään samasta isäntäresurssista kilpailevien kahden loislajin mahdollisia vuorovaikutussuhteita,
- suunnittelemaan kokeen, jolla määritetään näiden kahden lajin vuorovaikutussuhteiden luonne niiden kilpaillessa yhteisestä isännästä,
- Suorita konsensuskoe määrittääksesi lajinsisäisen ja lajienvälisen kilpailun vaikutukset Nasonia vitripenniksen ja Melittobia digitatan lisääntymistulokseen,
- Käytä tuloksena saatuja tietoja Lotka-Volterra-kilpailumallin parametrien estimoimiseksi,
- Suhteuta luokan tutkimustulokset kilpailun poissulkemisen periaatteeseen.
Ekologiset yhteisöt koostuvat elinympäristön kaikkien lajien populaatioista. Yhteisön rakenne määräytyy osittain yhteisön lajien välisten vuorovaikutusten dynamiikan perusteella. Kahden lajin väliset vuorovaikutukset voivat olla suoria tai epäsuoria (eli muiden lajien välittämiä).
Jopa yksinkertaisessa luonnonyhteisössä sadat eri kasvi- ja eläinlajit ovat vuorovaikutuksessa keskenään. Tästä monimuotoisuudesta huolimatta voimme kuitenkin tunnistaa vuorovaikutusluokkia, joilla on erilaisia vaikutuksia populaation kasvuun (taulukko 1). Luokat määritellään vuorovaikutuksessa oleviin lajeihin kohdistuvien vaikutusten suunnan mukaan.
Lajien välisten vuorovaikutusten lisäksi myös lajin sisäiset yksilöiden väliset vuorovaikutukset voivat olla tärkeitä yhteisön jäsentämisessä. Lajin sisäiset vuorovaikutukset voivat vaikuttaa lajin populaatiodynamiikkaan, joka puolestaan vaikuttaa lajien välisiin vuorovaikutuksiin. Lajinsisäistä kilpailua esiintyy, kun saman lajin tai populaation eri yksilöt kilpailevat resurssista. Nämä vuorovaikutukset voivat olla kovia, koska yksilöt tarvitsevat samoja rajallisia resursseja selviytyäkseen ja lisääntyäkseen. Kun eri lajit kilpailevat samasta ravinnosta, elinympäristöstä tai muusta ympäristöresurssista, sitä kutsutaan lajien väliseksi kilpailuksi. Nämä vuorovaikutussuhteet ovat yleensä jonkin verran lievempiä. Tämä johtuu siitä, että vaikka kahden lajin vaatimukset voivat olla samankaltaiset, ne eivät voi koskaan olla yhtä lähellä toisiaan kuin saman lajin yksilöillä.
| Taulukko 1. Samassa yhteisössä elävien kahden lajin välisten suorien vuorovaikutusten luokat | ||||||
| Vuorovaikutuksen nimi | Kontaktin tyyppi | Suora vaikutus lajiin #1 | Suora vaikutus lajiin #2 | Suhteen muut näkökohdat | ||
| Neutraali suhde | Kaksi lajia on yhteydessä toisiinsa vain epäsuorasti vuorovaikutuksen kautta muiden lajien kanssa. | 0 | 0 | Kullakin lajilla on neutraali suhde useimpiin lajeihin elinympäristössään. | ||
| Kommenssisuhde | Suhde, joka suoraan auttaa toista lajia, mutta joka ei vaikuta toiseen lajiin juurikaan, jos ollenkaan. | + | 0 | Kommensalismi, mutualismi ja parasitismi ovat kaikki symbioosin tapauksia. | ||
| Mutualismi | Hyötyjä virtaa molempiin suuntiin vuorovaikutuksessa olevien lajien välillä. | + | + | + | Parempi nähdä molemminpuolisena hyväksikäyttönä kuin viihtyisänä yhteistoimintana. | |
| Predation True Predators Grazers |
Predator hyökkää useiden saaliseläinten kimppuun ja ruokailee niitä, mutta ei asettaudu saaliseläimiin tai niiden päälle. | + | Saalis kuolee yleensä. Laiduntajien kanssa kasvi saattaa kuolla tai olla kuolematta. | |||
| Parasiitit Parasiitit Parasiitit Parasiitit | Parasiitti ruokkii yhden tai useamman isännän kudoksia ja asuu niissä tai niiden päällä ainakin osan elinkaarestaan. | + | Isäntä saattaa kuolla tai olla kuolematta vuorovaikutussuhteen tuloksena. | |||
| Sisälajikilpailu | Haitat voivat virrata molempiin suuntiin lajien välillä, tai ylivoimainen kilpailija voi jäädä suurelta osin ilman vaikutusta. | Yleisesti vähemmän voimakasta kuin saman lajin jäsenten keskinäinen kilpailu. | ||||
>0 tarkoittaa, että ei ole mitään välitöntä vaikutusta populaatiokasvustoon.
+ tarkoittaa positiivista vaikutusta; tarkoittaa negatiivista vaikutusta.
Tarkastellaan kuitenkin teoreettista tapausta, jossa kaksi lajia miehittää identtistä markkinarakoa. Gause (1934) tutki kahta protistilajia, jotka molemmat söivät samoja bakteerisoluja. Kun hän yhdisti ne samaan viljelyyn, toinen ajoi aina toisen sukupuuttoon. Monet muut kokeet ovat sittemmin tukeneet ”Gausen lakia”, jota nykyään kutsutaan nimellä kilpailullisen poissulkemisen periaate. Sen mukaan kaksi lajia, jotka käyttävät samoja resursseja, eivät voi elää rinnakkain loputtomiin tai ”täydelliset kilpailijat eivät voi elää rinnakkain” (Hardin 1960).
Monet kokeet ovat osoittaneet, että mitä enemmän kaksi lajia elinympäristössä eroaa toisistaan resurssien käytön suhteen, sitä todennäköisempää on, että ne voivat itse asiassa elää rinnakkain (Krebs 1994). Jopa kaksi lajia, joilla on paljon päällekkäisyyttä, voivat elää jonkin aikaa yhdessä, vaikka kilpailulliset vuorovaikutussuhteet usein tukahduttavat jommankumman tai molempien kasvuvauhdin. Ajan mittaan voi tapahtua mielenkiintoinen ilmiö, jota kutsutaan resurssien jakamiseksi. Kunkin lajin jäsenet voivat erikoistua jonkin samankaltaisten resurssien luokan osaan. Jos esimerkiksi molemmat lajit syövät omenoita, toinen voi syödä pieniä vihreitä hedelmiä ja toinen suurempia, kypsempiä hedelmiä.
Lotka-Volterra-malli kehitettiin, jotta ekologit voivat ennustaa mahdollisen lopputuloksen, kun kaksi lajia kilpailee samoista resursseista. Periaatteessa malli yrittää ottaa huomioon sen vaikutuksen, joka yhden lajin läsnäololla on toisen lajin populaation kasvuun suhteessa siihen kilpailuvaikutukseen, joka kahdella saman lajin edustajalla olisi toisiinsa nähden.
Yhtälö lajin 1 populaation kasvulle on:
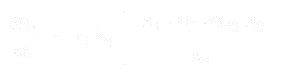
Ja lajin 2 osalta se on:
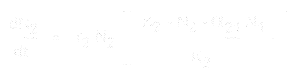
missä:
- * N1 ja N2 ovat lajien 1 ja 2 populaatiokoot,
* r1 ja r2 ovat näiden lajien luontaiset kasvunopeudet,
* K1 ja K2 ovat elinympäristön kantokapasiteetit kummallekin lajille,
* alpha12 ja alpha21 ovat yhden lajin vaikutukset toisen lajin populaation kasvuun. Tarkemmin sanottuna alpha12 on lajin 2 vaikutus lajin 1 kasvuun ja alpha21 on lajin 1 vaikutus lajin 2 kasvuun.
Jos kunkin yhtälön arvot tunnetaan (tai ne voidaan arvioida empiirisesti kokeen tuloksista), yhtälön avulla voidaan ennustaa mahdollisen kilpailun lopputulos (eli voivatko lajit elää rinnakkain vai syrjäyttääkö toinen lopulta toisen). K1:n, K2:n, a12:n ja a21:n arvojen avulla piirretään molempien lajien nollakasvun isokliinit (eli tilanteet, joissa dN1/dt tai dN2/dt on yhtä suuri kuin nolla) samaan kuvaajaan, ja tuloksena saatavia populaation kasvuvektoreiden summia (trajektorioita) käytetään kilpailun lopputuloksen määrittämiseen (kuva 1).

Kuva 1. Esimerkkikäyrästöt nollakasvun isoklinjoista, joissa lajit 1 ja 2 elävät rinnakkain (vasemmalla) ja laji 1 sulkee lajin 2 kilpailullisesti pois (oikealla).
Lotka-Volterran kilpailumalli kuvaa kahden lajin välisen kilpailun lopputulosta ekologisen ajan kuluessa. Koska yksi laji voi kilpailullisesti sulkea toisen lajin pois (kuva 1) ekologisessa ajassa, kilpailullisesti alempiarvoinen laji voi lisätä niiden ravintotyyppien valikoimaa, joita se syö selviytyäkseen. Lajien reaktio lajien väliseen kilpailuun evoluutioajassa on kuitenkin usein päinvastainen kuin ekologisessa ajassa. Kilpailijat erikoistuvat yleensä tiettyihin resurssityyppeihin. Tämä evoluutioajan kuluessa tapahtuva resurssien jakaminen johtaa itse asiassa siihen, että kahden lajin välinen kilpailu vähenee tai puuttuu kokonaan.
Kahden loispistiäislajin, Melittobia digitata ja Nasonia vitripennis, elämänhistoria on varsin samankaltainen, vaikka ne eivät olekaan erityisen läheistä sukua toisilleen. Molemmat lajit pystyvät käyttämään samaa isäntää, vaikka luonnossa ne käyttivätkin eri isäntiä. Melittobia-lajit ovat noin puolet pienempiä kuin Nasonia-lajit, mutta molemmat ovat melko pieniä ja ihmiselle täysin vaarattomia.
Niiden täydelliset elinkaaret ovat suhteellisen lyhyitä (2-4 viikkoa 25o C:ssa) ja myös melko samanlaisia (kuva 2). Naaraat munivat lukuisia munia isännän peitteen läpi. Munista kuoriutuu toukkia, jotka syövät isäntää, muuttuvat sitten poikasiksi ja lopulta metamorfoituvat aikuisiksi. Melitobbia digitatan aikuisilla naarailla voi olla joko normaalit tai kitukasvuiset siivet. Normaalisiipiset aikuiset hajaantuvat isännästä etsimään uusia ravintoresursseja. Lentokyvyttömät naaraat munivat munansa samaan isäntään, josta ne nousivat, tai hajaantuvat uuteen isäntään saman pesän sisällä (Freeman ja Ittyeipe 1976, Côsoli ja Vinson 2002).
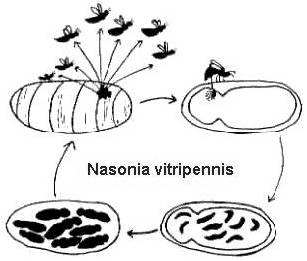
Kuvio 2. Lentokyvyttömät naaraat. Nasonia vitripenniksen elinkierto Neobellieria bullata -isännän poikasella (piirros Bethia King). Melittobia digitatan elinkierto on sama, vaikka yksilöt ovat kaikissa vaiheissa pienempiä.
Maailmanlaajuisesti tunnetaan noin 70 000 parasiittilajia (9 % kaikista hyönteisistä), mutta arvioita niiden määrästä on jopa 800 000 (Strand 2002). Kuvatuista lajeista noin 80 % kuuluu Hymenoptera-lahkoon, kuten myös ne kaksi lajia, joita käytät tässä laboratoriossa. Luonnossa isäntiä voi loistaa useampi kuin yksi parasiittilaji, jotka kaikki kilpailevat samasta resurssista (Stand 2002). Koska isäntälaji voi itse olla toisen lajin loinen, tämän kilpailun ekologiset vaikutukset yhteisön rakenteeseen voivat olla hyvin monimutkaisia. Esimerkiksi Swaine-mäntysahakärpäset (Neodiprion swainei Midd.), jotka voivat hyökätä, tuhota ja tappaa suuria mäntykasvustoja Pohjois-Amerikassa, käyttävät isäntänään 11:tä erilaista loispistiäislajia. Näyttää siltä, että useimmat näistä lajeista elävät rinnakkain, koska ne jakavat isännän resurssit keskenään loisimalla isännän elinkaaren eri vaiheissa tai isäntäpopulaation eri osissa (Price 1972). Yksi sääkärpäskokkeja vastaan hyökkäävistä loispistiäisten killan jäsenistä on alueelle tuotu laji (Pleolophus basizonus). Se on ylivoimainen kilpailija, ja sen läsnäolo määrää joidenkin kilpailijalajien runsauden (Price 1970). Vaikka kilpailu voi vaikuttaa loispistiäisyhteisöjen rakenteeseen, isäntäresurssien alueellinen ja ajallinen vaihtelu voi olla vielä tärkeämpää loispistiäislajien lajirunsauden määrittelyssä, vaikka loispistiäisten välinen kilpailu olisikin olemassa (Hawkins 2000).
Parasiitoideja, joiden isännät ovat tärkeitä viljelykasvien tai metsäpuiden tuholaisia, vapautetaan joskus tarkoituksellisesti tuholaispopulaatioiden biologiseksi torjumiseksi. Jos useampi kuin yksi parasiitoidilaji hyökkää samaa isäntää vastaan, on tärkeää ymmärtää kilpailun luonne ennen levittämistä. Jos eri loispistiäislajit pystyvät elämään rinnakkain resurssien jakamisen avulla, tuholaisisännän torjunta voidaan ehkä parhaiten saavuttaa istuttamalla joitakin tai kaikkia loispistiäislajeja. Toisaalta, jos kilpailijat rajoittavat toistensa populaatioita kilpailullisen vuorovaikutuksensa vuoksi, tuholaislajin parasitoidi voidaan torjua parhaiten vapauttamalla parasitoidilajeista tehokkain laji yksinään (Amarasekare 2000).
Materiaalit ja menetelmät:
Katsaus tiedonkeruu- ja analyysimenetelmiin:
Lab 1:
Taulukossa 1 käsitellyt vuorovaikutusluokat voivat tuntua melko suoraviivaisilta, kun niistä vain lukee. Mutta jos havainnoisit kahta tuntematonta eläintä vuorovaikutuksessa, miten päättäisit, mitä ”etikettiä” käyttäisit? Voisitko ennustaa vuorovaikutuksen lopputuloksen? Miten voisit testata ennustuksesi?
Tässä laboratoriotutkimuksessa esiteltävät kaksi loispistiäistä näyttävät miehittävän samanlaisia markkinarakoja. Olemme kiinnostuneita näiden kahden lajin välisistä vuorovaikutuksista. Lisäksi haluamme pystyä kvantifioimaan toisen lajin vaikutusta toiseen lajiin.
Kummankin ryhmän tulisi:
- – keskustella ja luetella kaikki mahdolliset koeyhdistelmät, jotka voitaisiin perustaa kahden loisampiaisen, Melittobia digitatan ja Nasonia vitripenniksen, ja yhden isännän, Neobellierrian, välille,
– ennustaa, mikä olisi mielestänne kunkin mahdollisen vuorovaikutuksen tulos,
– yksilöi ja luettele muuttujat, joita manipuloisit kokeessasi,
– yksilöi ja luettele muuttujat, jotka pitäisit kokeessasi vakiona,
– yksilöi ja luettele riippuvaiset muuttujat, joita haluaisit mitata, jotta voisit kvantifioida kummankin lajin vaikutuksen toisiinsa.
Jokainen ryhmä jakaa koesuunnitelmansa muun luokan kanssa. Luokkana kehitämme yhdessä yksimielisen koesuunnitelman. Konsensukseen perustuvan koesuunnitelman perusteella kunkin henkilön tulisi perustaa yksi toistoviljelmä kutakin käsittelyä varten. Katso ”Parasitoidien käsittely” jäljempänä. Kokoamme koko luokan tiedot yhteen analysointia varten.
Viikoittaiset tarkastukset:
Joka viikko jokaisen henkilön tulisi tarkastaa viljelmänsä nähdäkseen, onko aikuisia ampiaisia syntynyt. Sinun tulisi kirjata päivämäärä, jolloin näet ensimmäisen kerran aikuisia ampiaisia kunkin viljelmän kohdalla. Nasonia-viljelmät olisi pakastettava 21 päivän kuluttua niiden perustamisesta. Melittobia- ja sekalajiviljelmät olisi pakastettava 42 päivän kuluttua.
Laboratorio 2 (6 viikkoa laboratorion 1 jälkeen):
Kunkin henkilön on laskettava kussakin toistoviljelmässä tuotettujen jälkeläisten lukumäärä. Syötä tietosi taulukkolaskentaohjelmaan, jotta luokan tiedot voidaan yhdistää. Käytä yhdistettyjä tietoja Lotka-Volterra-kilpailumallin parametrien estimointiin ja kilpailun vaikutusten tilastolliseen analysointiin.
Keskustelkaa ryhmissänne siitä, miten aineistoa voidaan käyttää Lotka-Volterran kilpailumallin parametrien kvantifiointiin (ks. ”Lokta-Volterran kilpailumallin kvantifiointi” jäljempänä). Keskustele myös siitä, mitä tietyt käsittelyvertailut voivat kertoa meille lajinsisäisen ja lajienvälisen kilpailun suhteellisesta merkityksestä näissä kahdessa loispistiäislajissa (ks. ”Ohjeita tietojen analysointiin” jäljempänä).
Lasitoidien käsittely:
Kummankin parasiitoidin, Melittobia digitatan ja Nasonia vitripennisin, aikuiset yksilöt ovat hyvin ”käyttäjäystävällisiä”. Vaikka naarailla on normaalit siivet ja ne voivat lentää, ne eivät tee sitä helposti. Ne ovat kuitenkin negatiivisesti geotaksisia (eli ne liikkuvat ylöspäin, poispäin painovoimasta). Kun muutamaa naarasta ravistellaan vaakasuoralle pinnalle ja peitetään sen jälkeen ylösalaisin käännetty lasipullo, ne kiipeävät helposti pulloon ja kiipeävät sen sivuja ylöspäin. Kun ampiaisia on pullossa, voit helposti lisätä isäntäkukan ja tukkia pullon tiiviisti puuvillalla. Tällä tavoin voidaan käsitellä tehokkaasti suuria määriä yksilöitä. Aikuisia ampiaisia voidaan käsitellä myös lyhyillä putkisiivoojilla, joihin ampiaiset tarttuvat väliaikaisesti.
Lotka-Volterran kilpailumallin kvantifiointi:
Lotka-Volterra-kilpailumalli kuvattiin ja määriteltiin yhtälöineen johdannossa. Kuten siellä todettiin, jos kunkin yhtälön arvot voidaan arvioida empiirisesti kokeen tuloksista, yhtälön avulla voidaan ennustaa kilpailun mahdollinen lopputulos (ts. se, elävätkö kaksi lajia rinnakkain vai syrjäyttääkö toinen laji lopulta toisen). K1:n, K2:n, alfa12:n ja alfa21:n arvoja käytetään piirtämään molempien lajien nollakasvun isokliinit (eli missä dN1/dt tai dN2/dt on nolla) samaan kuvaajaan, ja tuloksena saatavia populaation kasvuvektoreiden summia (lentoratoja) käytetään kilpailun lopputuloksen määrittämiseen.
Kokeellisen suunnitelmamme perusteella meidän on määritettävä näiden parametrien arvot. Muistutetaan, että populaation kantokyky on suurin yksilömäärä, joka voi selviytyä elinympäristössä. Yksinkertaisuuden vuoksi olemme tässä kokeessa määritelleet loispistiäisten elinympäristöksi yhden isännän. Todellisuudessa elinympäristö sisältäisi todennäköisesti useamman kuin yhden mutakärpäsen pesän tai isokärpäsen pupariumin, joten potentiaalisesti hyödynnettävissä olevia isäntiä olisi monia. Kahden lajin kantokyvyn määrittämiseksi meidän on tiedettävä tietyn lajin jälkeläisten enimmäismäärä, joka voidaan tuottaa yhdellä isännällä, kun vain kyseinen laji on läsnä. Minkä käsittelyn tietoja käytettäisiin Melittobian ja Nasonian kantokyvyn arvioimiseksi? (Muista, että kantokapasiteetin ollessa saavutettu kaikki isännän resurssit käytetään.)
Kilpailukertoimien (alpha12 ja alpha21) arvioiminen on hieman monimutkaisempaa. Muistetaan, että lajin 1 populaation kasvun yhtälö on:
Kun kaikki isäntäresurssi on loislääkkeiden käytössä, populaatio ei voi enää kasvaa. Toisin sanoen dN1 /dt = 0. Tämä ehto toteutuu, kun K1 – N1 – alpha12*N2 = 0. Löytääksemme alpha12 meidän on ratkaistava se (eli tehtävä hieman algebraa) ja sitten korvattava arvot K1:lle, N1:lle ja N2:lle. Edellä kuvattiin, miten kantavuudet löydetään. Jos oletetaan, että Melittobia on ”laji 1”, käytetään sen kantokykyä K1:lle. Lajien välisessä kilpailussa syntyvien Melittobian ja Nasonian jälkeläisten määrä on N1 ja N2. Minkä käsittelyn tietoja käytettäisiin N1:n ja N2:n arvioimiseksi?
Samaa lähestymistapaa, jota käytit alfa12:n laskemiseen, voidaan käyttää myös alfa21:n laskemiseen.
Nyt kun olet laskenut kaikki parametriarvot, voit käyttää näitä arvoja piirtääksesi nollakasvun isokliinit ja ennustaa Melittobian ja Nasonian välisen kilpailun tuloksen.
Ohjeita tietojen analysointiin:
Voidaan käyttää eri käsittelyjen välisiä vertailuja lajinsisäisen ja lajien välisen kilpailun suhteellisen merkityksen tutkimiseen. Määritä ensin, minkä tyyppistä kilpailua, lajinsisäistä tai lajien välistä, esiintyy kussakin käsittelyssä, jos sitä esiintyy. Kun olet tehnyt tämän, mieti kaikkia käsittelyparien välisiä vertailuja. Mitä kukin vertailu kertoo meille? Voi olla hyödyllistä laatia taulukko, jossa luetellaan vertailut ja niiden merkitys. Koska kaikki vertailut koskevat kahta hoitoa, niitä voidaan analysoida tilastollisesti t-testien avulla.
Kysymyksiä jatkopohdintaa ja keskustelua varten:
- Laskemiesi parametriarvojen perusteella, jotka laskit Lotka-Volterra-kilpailumallille, mikä on kahden lajin välisen kilpailun ennustettu tulos? Saavutettiinko ennustettu tulos jokaisessa lajien välisen kilpailun replikaatiossa? Jos ei, niin miksi ei?
- ”Gausen laki” sanoo, että kilpailijat, jotka jakavat täsmälleen samat resurssit samalla tavalla, eivät voi elää rinnakkain. Tämä tarkoittaa sitä, että se laji, joka käyttää kiisteltyä resurssia tehokkaimmin, hävittää lopulta toisen lajin kyseisestä paikasta. Näyttääkö Gausen laki pätevän Melittobian ja Nasonian väliseen vuorovaikutukseen? Miksi vai miksi ei?
- Jos nämä kaksi lajia käyttäisivät samaa isäntää luonnossa, miten resurssien jakaminen mahdollistaisi niiden rinnakkaiselon?
- Kokeesi tulosten perusteella, miksi nämä kaksi lajia eivät käytä samaa isäntää luonnossa?
- Kannatuskapasiteetin ja kilpailukertoimien arvioidut arvot huomioon ottaen ennusta Melittobian ja Nasonian välisen kilpailun lopputulos käyttäen Lotka-Volterran kilpailumallia Populuksessa (ks. Viitteet ja linkit). Vaikuttavatko alkuperäiset populaatiokoot tai populaation kasvunopeudet ennustettuun kilpailutulokseen? Jos vaikuttaa, miten? Miten nämä arvot vaikuttavat tasapainon saavuttamiseen kuluvaan aikaan?
- Kannatuskapasiteetit ja kilpailukertoimet ovat vain arvioita. Mitkä tekijät voivat vaikuttaa näiden kahden lajin kantokapasiteettiin ja kilpailukertoimeen?
- Jos näissä lajeissa esiintyy lajien välistä kilpailua, miten voisimme määrittää, mikä kilpailumekanismi (häiriö vai hyväksikäyttö) on kyseessä?
*** Huomautus: Vastaukset moniin näistä kysymyksistä ja lukuisat muut kommentit, jotka kirjoittaja on esittänyt, löytyvät asiakirjasta ”HUOMAUTUKSIA OPETTAJALLE: Comments On Questions for Further Thought” -sivulla.
Viitteet ja linkit:
- Abe, J., Y. Kamimura, N. Kondo ja M. Shimada. 2003. Äärimmäisen naaraspuolinen sukupuolisuhde ja tappava uroksen ja uroksen välinen taistelu loispistiäisessä, Melittobia australica (Eulophidae). Behavioural Ecology 14: 34-39.
- Amarasekare, P. 2000. Kilpailevien loispistiäisten rinnakkaiselo hajanaisesti jakautuneella isännällä: paikalliset vs. alueelliset mekanismit. Ecology 81: 1286-96.
- Cooperband M. F., R. W. Matthews, S. B. Vinson. 2003. Melittobia digitatan (Hymenoptera: Eulophidae) lisääntymisbiologiaan vaikuttavat tekijät ja Hamiltonin paikallisen parituskilpailun teorian sukupuolisuhde-ennusteiden toteutumattomuus. Entomologia Experimentalis et Applicata 109: 1-12.
- Côsoli, F. L. ja S. B. Vinson. 2002. Melittobia digitatan pesäkoko, kehitys ja siipimorfien erilaistuminen. Entomologia Experimentalis et Applicata 102: 135-143.
- Freeman, B. E., and K. Ittyeipe. 1976. Field studies on the cumulative response of Melittobia sp. Hawaiiensis complex) (Eulophidae) to varying host densities. Journal of Animal Ecology 45: 415-423.
- Gause, G. F. 1934. Taistelu olemassaolosta. Williams and Wilkins, Baltimore.
- Harden, G. 1960. Kilpailun poissulkemisen periaate. Science 131: 1292-1297.
- Hawkins, B. A. 2000. Lajien rinnakkaiselo loispistiäisyhteisöissä: onko kilpailulla väliä? Sivut 198-213 teoksessa M. E. Hochberg ja A. R. Ives (toim.). Parasitoid Population Biology. Princeton University Press, Princeton, New Jersey.
- King, B. 2000. Nasonia vitripennis, jalokiviampiainen. Tieteen ja biologian käsitteiden havainnollistaminen hyönteisen avulla. www.bios.niu.edu/bking/nasonia.htm
- Krebs, C. J. 1994. Ecology. Levinneisyyden ja runsauden kokeellinen analysointi. Neljäs painos. HarperCollins College Publishers, New York, NY.
- Mackauer, M., B. Bai, A. Chow ja T. Danyk. 1992. Epäsymmetrinen toukkakilpailu kahden yksinäisen loispistiäislajin välillä: superparasiittien vaikutus. Ecological Entomology 17: 233-236.
- Price, P. 1970. Samaa isäntää käyttävät loispistiäiset: koko- ja muotoerojen adaptiivinen luonne. Ecology 53: 445-454.
- Price, P. 1972. Quebecissä sijaitsevan sahakärpäsen parasiitoidien rinnakkaiselon mahdollistavat ominaisuudet. Ecology 51: 190-195.
- Strand, M. R. 2002. Toukkavaiheen parasiitoidien ja niiden isäntien väliset vuorovaikutukset. Sivut 129-152 teoksessa E. E. Lewis, J. F. Campbell ja M. V. K. Sukhdeo (toim.). The Behavioural Ecology of Parasites. CABI Publishing, New York.
- Silva-Torres, C., ja R. W. Matthews. 2003. Neobellieria bullata (Parker) (Diptera: Sarcophagidae) -yksilön (Diptera: Sarcophagidae) pupariaan loisivan Melittobia australica Girault’n ja M. digitata Dahmsin (Hymenoptera: Eulophidae) kehitys. Neotropical Entomology 32: 645-651.
- Werren, J. H. 1983. Sukupuolisuhteen evoluutio paikallisen parituskilpailun vallitessa loisevassa ampiaisessa. Evolution 37:116-124.
- Werren, J. H. 1984. Pesäkoon ja sukupuolisuhteen säätely parasiittiampiaisessa Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae). Netherlands Journal of Zoology 34: 123-143.
Tietoa luonnonhistoriasta ja elinympäristöistä löytyy osoitteesta:
- www.wowbugs.com for Melittobia,
www.bios.niu.edu/bking/nasonia.htm for Nasonia,
www.rochester.edu/College/BIO/labs/WerrenLab/nasonia/ for Nasonia.
Populus voidaan ladata osoitteesta www.cbs.umn.edu/populus.
Tools for Assessment of Student Learning Outcomes:
Arviointi on toteutettu eri tavoin eri oppilaitoksissa, jotka ovat käyttäneet tätä harjoitusta. Kaikissa tapauksissa opiskelijat on arvioitu tieteellisen tutkielman perusteella, jonka kukin opiskelija on kirjoittanut yksin tai opiskelijat ovat kirjoittaneet ryhmässä. Joissakin tapauksissa opiskelijat on arvioitu sekä ensimmäisen että toisen luonnoksen perusteella.
Pisteytysohjeet vaihtelevat opettajan mukaan. Alla on esimerkki Morehouse Collegessa käytetystä arvostelurubriikista, joka koskee ”tulosyhteenvetoa”, jossa on kaikki tieteellisen tutkielman osat menetelmiä lukuun ottamatta. Tässä arviointikriteeristössä ”yleisö” koskee opiskelijan tekemää tarkoituksenmukaisen yleisön valintaa. Opiskelijoiden odotetaan kirjoittavan raporttinsa ikään kuin se olisi tieteellinen artikkeli. Sopiva yleisö on siis vertaisryhmä, joka ei ole suorittanut koetta mutta joka on tieteellisesti lukutaitoinen. Opettajalle tai muille luokan opiskelijoille kirjoitetuilla raporteilla ei ole sopivaa yleisöä. ”Muotoilulla” tarkoitetaan raportin yleistä järjestämistä osiin, joilla on samansuuntainen organisaatio ja jotka rakentuvat toistensa päälle. Esimerkiksi Keskustelussa olisi arvioitava Tuloksissa raportoituja tuloksia ja asetettava ne laajempaan yhteyteen. Keskustelussa olisi myös käsiteltävä johdannossa esitettyä hypoteesia.
| Tulosten yhteenvedon arviointi (50 pistettä mahdollista) | |
| Esittely ja otsikkosivu (10 pistettä) | _____ |
| Tulokset (10 pistettä). pistettä) | _____ |
| Keskustelu ja johtopäätökset (10 pistettä) | _____ |
| Kirjallisuuden käyttö ja viittaukset (10 pistettä) | _____ |
| Format, Yleisö (10 pistettä) | _____ |
Tutustumisvaiheessa opiskelijat toimittavat kaikki luokassa laaditut taulukot, vastaukset keskustelukysymyksiin ja viikoittaiset muistiinpanot tutkimuksen edistymisestä. Yksilöllisten tieteellisten töiden lisäksi opiskelijat voisivat esitellä kokeen tulokset ryhmätyönä tieteellisissä töissä, ryhmätyönä suullisissa esityksissä tai ryhmätyönä posteriesityksinä. Koska kaikki oppilaat suorittavat kuitenkin saman kokeen ja esittävät näin ollen samat tulokset, yksilölliset tai ryhmäkohtaiset tieteelliset paperit olisivat tehokkaimpia.
Työkaluja tämän kokeen formatiiviseen arviointiin:
Morehouse Collegen ekologian kurssilla jokaista opiskelijaa pyydetään arvioimaan kutakin laboratorioharjoitusta 10 pisteen asteikolla sen suhteen, kuinka tehokkaaksi kukin tutkimus koetaan vahvistamaan heidän tietämystään ja ymmärrystään ekologian luentokurssilla käsitellyistä aiheista. Lisäksi opiskelijoilta kysytään, mitkä harjoitukset olivat vähiten ja mitkä eniten miellyttäviä ja mitkä lisäsivät eniten heidän ymmärrystään tieteellisestä menetelmästä. Näitä tietoja käytetään seuraavien lukukausien harjoitusten valinnassa ja muokkaamisessa.
Emoryn yliopiston ja Radfordin yliopiston ekologian kursseilla jokaiselta opiskelijalta kysytään, mistä harjoituksista hän piti eniten ja mistä vähiten. Opiskelijat antavat joskus kommentteja siitä, miksi he arvioivat harjoitukset niin kuin he arvioivat. Myös tätä tietoa käytetään seuraavien lukukausien harjoitusten muokkaamiseen.
Laaja keskustelu arvioinnista on tämän sivuston Opetusosiossa.
.