Detailed Description of the Experiment
- Introduction
- Materials and Methods
- Questions til yderligere overvejelser og diskussion
- Referencer og links
- Værktøjer til vurdering af elevernes læringsresultater
- Værktøjer til formativ evaluering af dette forsøg
Indledning:
I denne undersøgelse vil vi undersøge virkningerne af konkurrence om ressourcer på reproduktionsoutput inden for og mellem to arter af parasitoid hvepse.
Laboratoriemål:
Efter afslutningen af dette forsøg vil eleverne være i stand til at…
- Beskrive livscyklusen for Nasonia vitripennis og Melittobia digitata,
- Forklare de mulige interaktioner mellem to parasitarter, der konkurrerer om den samme værtsressource,
- Udforme et eksperiment for at bestemme arten af interaktionen mellem disse to arter, når de konkurrerer om en fælles vært,
- Udføre et konsensusforsøg for at bestemme virkningerne af intraspecifik og interspecifik konkurrence på reproduktionsoutput hos Nasonia vitripennis og Melittobia digitata,
- Bruge de resulterende data til at estimere parametrene for Lotka-Volterra-konkurrencemodellen,
- Sætte resultaterne af klasseforskningen i relation til princippet om udelukkelse af konkurrence.
Økologiske samfund er sammensat af populationer af alle arter i et levested. Strukturen i et samfund vil til dels være bestemt af dynamikken i samspillet mellem arterne i samfundet. Interaktioner mellem to arter kan være direkte eller indirekte (dvs. formidlet gennem andre arter).
I selv et simpelt naturligt samfund er der hundredvis af forskellige arter af planter og dyr, der interagerer med hinanden. På trods af denne mangfoldighed kan vi imidlertid identificere kategorier af interaktioner, som har forskellige virkninger på populationsvæksten (tabel 1). Kategorierne er defineret efter retningen af virkningerne på de interagerende arter.
Ud over interaktioner mellem arter kan interaktioner mellem individer inden for en art også være vigtige for struktureringen af et samfund. Interaktioner inden for arterne kan påvirke artens populationsdynamik, hvilket igen vil påvirke interaktionerne mellem arterne. Intraspecifik konkurrence opstår, når forskellige individer af den samme art eller population konkurrerer om en ressource. Disse interaktioner kan være voldsomme, fordi individerne har brug for de samme begrænsede ressourcer for at overleve og reproducere sig. Når forskellige arter kæmper om den samme føde, det samme levested eller en anden miljøressource, kaldes det interspecifik konkurrence. Disse interaktioner er typisk noget mindre intense. Det skyldes, at selv om kravene hos to arter kan være ens, kan de aldrig være så tæt på hinanden, som de er for individer af samme art.
| Tabel 1. Kategorier af direkte interaktioner mellem to arter i samme samfund | ||||||
| Navn på interaktionen | Type af kontakt | Direkte effekt på art #1 | Direkte virkning på art nr. 2 | Andre aspekter af forholdet | ||
| Neutralt forhold | To arter er kun indirekte forbundet gennem interaktioner med andre arter. | 0 | 0 | Hver art har et neutralt forhold til de fleste arter i dens levested. | ||
| Kommensalisme | Et forhold, der direkte hjælper den ene art, men som ikke påvirker den anden meget, hvis overhovedet. | + | 0 | Kommensalisme, mutualisme og parasitisme er alle tilfælde af symbiose. | ||
| Mutualisme | Fordelene flyder begge veje mellem de arter, der interagerer. | + | + | + | Det kan bedre ses som tovejsudnyttelse end som et hyggeligt samarbejde. | |
| Predation Egte rovdyr Grazers |
Predator angriber og æder en række byttedyr, men tager ikke ophold i eller på dem. | + | Byttet dør som regel. Med græsædere dør planten måske eller måske ikke. | |||
| Parasitisme Parasitter Parasitoider | Parasitten lever af væv fra en eller flere værter og opholder sig i eller på dem i mindst en del af deres livscyklus. | + | Værten dør måske eller måske ikke som følge af samspillet. | |||
| Interspecifik konkurrence | Ulemper kan flyde begge veje mellem arterne, eller den overlegne konkurrent kan være stort set upåvirket. | Generelt mindre intens end konkurrence mellem medlemmer af samme art. | ||||
0 betyder ingen direkte effekt på populationsvæksten.
+ betyder positiv effekt; betyder negativ effekt.
Og tænk dog på det teoretiske tilfælde med to arter, der besætter den samme niche. Gause (1934) undersøgte to protistarter, der begge ernærede sig af de samme bakterieceller. Når han kombinerede dem i en enkelt kultur, drev den ene altid den anden til udryddelse. Mange andre eksperimenter har siden da understøttet “Gauses lov”, som nu kaldes princippet om konkurrencemæssig udelukkelse. Den fastslår, at to arter, der udnytter identiske ressourcer, ikke kan eksistere side om side på ubestemt tid, eller at “fuldstændige konkurrenter ikke kan eksistere side om side” (Hardin 1960).
Flere eksperimenter har vist, at jo mere to arter i et levested adskiller sig fra hinanden i deres ressourceudnyttelse, jo mere sandsynligt er det, at de rent faktisk kan eksistere side om side (Krebs 1994). Selv to arter med stor overlapning kan leve sammen i nogen tid, selv om konkurrencemæssige interaktioner ofte undertrykker den ene eller begge arters vækstrate. Over tid kan der opstå et interessant fænomen kaldet ressourcefordeling. Medlemmer af hver art kan komme til at specialisere sig i en underopdeling af en eller anden kategori af lignende ressourcer. Hvis begge arter f.eks. lever af æbler, kan den ene leve af små grønne frugter og den anden af større, mere modne frugter.
Lotka-Volterra-modellen blev udviklet for at gøre det muligt for økologer at forudsige det potentielle resultat, når to arter konkurrerer om de samme ressourcer. Grundlæggende forsøger modellen at tage højde for den virkning, som tilstedeværelsen af den ene art vil have på den anden arts populationsvækst, i forhold til den konkurrerende virkning, som to medlemmer af samme art ville have på hinanden.
Ligningen for populationsvæksten for art 1 er:
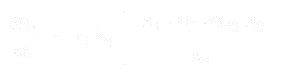
Og for art 2 er den:
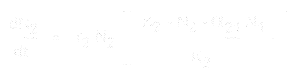
hvor:
- * N1 og N2 er populationsstørrelserne for art 1 og 2,
* r1 og r2 er de iboende vækstrater for disse arter,
* K1 og K2 er levestedets bæreevne for hver art,
* alpha12 og alpha21 er den ene arts virkninger på den anden arts populationsvækst. Nærmere bestemt er alpha12 den virkning, som art 2 har på væksten af art 1, og alpha21 er den virkning, som art 1 har på væksten af art 2.
Hvis værdierne for hver ligning er kendt (eller kan estimeres empirisk ud fra resultaterne af et eksperiment), kan ligningen bruges til at forudsige det potentielle resultat af en konkurrence (dvs. om de kan sameksistere, eller om den ene i sidste ende vil udelukke den anden). Værdierne for K1, K2, a12 og a21 bruges til at plotte isoklinerne for nulvækst (dvs. hvor dN1/dt eller dN2/dt er lig nul) for begge arter på den samme graf, og de resulterende summer af populationsvækstvektorer (trajektorier) bruges til at bestemme resultatet af konkurrencen (figur 1).

Figur 1. Eksempelgrafer af isoklinier med nulvækst, for hvilke art 1 og art 2 eksisterer side om side (til venstre), og art 1 udelukker art 2 i konkurrence med art 1 (til højre).
Lotka-Volterra-konkurrencemodellen beskriver resultatet af konkurrencen mellem to arter over økologisk tid. Fordi en art kan udelukke en anden art konkurrencemæssigt (figur 1) i økologisk tid, kan den konkurrencemæssigt underlegne art øge det udvalg af fødevaretyper, som den spiser, for at overleve. Arternes reaktion på interspecifik konkurrence i evolutionær tid er imidlertid ofte det modsatte af, hvad der sker i økologisk tid. Konkurrenterne vil generelt specialisere sig på bestemte ressourcetyper. Denne ressourceopdeling, der sker i løbet af evolutionær tid, resulterer faktisk i mindre eller ingen konkurrence mellem de to arter.
Selv om de ikke er særligt nært beslægtede med hinanden, er livshistorien for to parasitoid hvepsearter, Melittobia digitata og Nasonia vitripennis, ret ens. Begge arter er i stand til at bruge den samme vært, selv om de i naturen brugte forskellige værter. Melittobia er ca. halvt så store som Nasonia, men begge er ret små og helt ufarlige for mennesker.
Deres komplette livscyklus er relativt kort (2-4 uger ved 25o C) og er også ret ens (figur 2). Hunnerne lægger mange æg gennem værtsdækket. Æggene klækkes og bliver til larver, der æder værten, derefter forvandles de til pupper og til sidst metamorpherer til et voksent stadium. Hos Melitobbia digitata kan de voksne hunner have enten normale eller forkrøblede vinger. De normale voksne med normale vinger spreder sig fra værten for at søge efter nye fødekilder. De vingeløse hunner lægger deres æg på den samme vært, som de kom ud af, eller de spreder sig til en ny vært inden for samme rede (Freeman og Ittyeipe 1976, Côsoli og Vinson 2002).
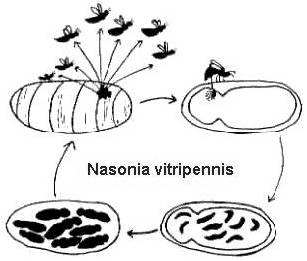
Figur 2. Livscyklus af Nasonia vitripennis på en Neobellieria bullata-værtspuppe (tegning af Bethia King). Livscyklusen hos Melittobia digitata er den samme, selv om individerne på alle stadier er mindre.
Der er ca. 70.000 kendte arter af parasitoider på verdensplan (9 % af alle insekter), men antallet skønnes at være helt op til 800.000 (Strand 2002). Af de beskrevne arter tilhører ca. 80 % hymenoptera-ordenen, ligesom de to arter, du bruger i denne opgave. I naturen kan værter blive parasiteret af mere end én art af parasitoid, som alle konkurrerer om den samme ressource (Stand 2002). Da værtsarten selv kan være en parasit på en anden art, kan de økologiske virkninger af denne konkurrence på samfundsstrukturen være meget komplekse. F.eks. er Swaine jack pine sawflies (Neodiprion swainei Midd.), som kan angribe, afblomstre og dræbe store bevoksninger af jack pines i Nordamerika, vært for 11 forskellige arter af parasitoid hvepse. Det ser ud til, at de fleste af disse arter eksisterer side om side, fordi de fordeler værtsressourcen mellem sig ved at parasitere forskellige stadier af værtens livscyklus eller forskellige segmenter af værtspopulationen (Price 1972). Et medlem af den parasitoidegilde, der angriber savfluekokoner, er en indført art (Pleolophus basizonus). Den er en overlegen konkurrent, og dens tilstedeværelse bestemmer mængden af nogle af dens konkurrerende arter (Price 1970). Selv om konkurrence kan påvirke parasitoidernes samfundsstruktur, kan den rumlige og tidsmæssige variation i værtsressourcerne være endnu vigtigere for bestemmelsen af parasitoidernes artsrigdom, selv i tilfælde af interspecifik konkurrence mellem parasitoiderne (Hawkins 2000).
Parasitoider, hvis værter er vigtige skadedyr på afgrøder eller skovtræer, udsættes undertiden med vilje som biologisk kontrol af skadedyrsbestande. Når mere end én parasitoidart angriber den samme vært, er det vigtigt at forstå arten af denne konkurrence, før der foretages udsætninger. Hvis de forskellige parasitoidarter er i stand til at eksistere side om side ved hjælp af ressourcefordeling, kan bekæmpelse af skadedyrsværten bedst opnås ved at indføre nogle eller alle parasitoidarterne. Hvis konkurrenterne på den anden side begrænser hinandens populationer på grund af deres konkurrencemæssige interaktioner, kan der opnås maksimal bekæmpelse af skadegørerarten ved at udsætte den mest effektive af parasitoidarterne alene (Amarasekare 2000).
Materialer og metoder:
Overblik over dataindsamling og analysemetoder:
Laboratorium 1:
De kategorier af interaktioner, der diskuteres i tabel 1, kan virke ret ligetil, når man blot læser om dem. Men hvis du skulle observere to ukendte dyr, der interagerer, hvordan ville du så beslutte, hvilken “etiket” du skulle anvende? Kunne du forudsige resultatet af interaktionen? Hvordan kunne du teste din forudsigelse?
De to snyltehvepse, der præsenteres i denne laboratorieundersøgelse, synes at besætte lignende nicher. Vi er interesseret i interaktionerne mellem de to arter. Desuden ønsker vi at kunne kvantificere den ene arts virkning på den anden.
Hver gruppe skal:
- – diskutere og opregne alle de mulige forsøgskombinationer, der kunne opstilles, og som involverer to snyltehvepse, Melittobia digitata og Nasonia vitripennis, og en enkelt vært, Neobellierria,
– forudsige, hvad I tror, der kan blive resultatet for hver mulig interaktion,
– identificere og opregne de variabler, som du vil manipulere i dit forsøg,
– identificere og opregne de variabler, som du vil holde konstante i dit forsøg,
– identificere og opregne de afhængige variabler, som du vil måle for at kvantificere hver arts virkning på den anden.
Hver gruppe skal dele deres forsøgsplan med resten af klassen. Sammen som klasse vil vi udvikle en fælles forsøgsplan. På grundlag af konsensusforsøgsdesignet skal hver person opstille en gentagelseskultur for hver behandling. Se “Håndtering af parasitoider” nedenfor. Vi samler dataene fra hele klassen med henblik på analyse.
Vejlige kontroller:
Hver uge skal hver person undersøge sine kulturer for at se, om der er kommet voksne hvepse frem. De skal notere den dato, hvor de første gang ser voksne hvepse fremkomme for hver kultur. Nasonia-kulturer skal fryses 21 dage efter, at de er blevet etableret. Melittobia- og blandingsarterkulturer bør fryses efter 42 dage.
Laboratorium 2 (6 uger efter laboratorium 1):
Hver person skal tælle antallet af afkom, der er produceret i hver replikatkultur. Indtast jeres data i et regneark, så dataene for klassen kan samles i et regneark. Brug de sammenlagte data til at estimere parametrene for Lotka-Volterra-konkurrencemodellen og til statistisk analyse af virkningerne af konkurrencen.
Diskuter i jeres grupper, hvordan dataene kan bruges til at kvantificere parametrene for Lotka-Volterra-konkurrencemodellen (se “Kvantificering af Lokta-Volterra-konkurrencemodellen” nedenfor). Diskuter også, hvad særlige behandlingssammenligninger kan fortælle os om den relative betydning af intraspecifik og interspecifik konkurrence hos disse to arter af parasitoider (se “Retningslinjer for dataanalyse” nedenfor).
Håndtering af parasitoider:
Voksne af begge parasitoider, Melittobia digitata og Nasonia vitripennis, er meget “brugervenlige”. Selv om hunnerne har normale vinger og kan flyve, gør de det ikke let. De er dog negativt geotaksiske (dvs. de bevæger sig opad, væk fra tyngdekraften). Når nogle få hunner fra en kultur rystes ud på en vandret overflade og derefter dækkes med en omvendt glaskolbe, klatrer de let ind i kolben og op ad siderne. Når man først har hvepse i et glasglas, kan man let tilføje en værtspuppe og derefter lukke glasset tæt til med vat. Et stort antal individer kan håndteres effektivt på denne måde. De voksne hvepse kan også manipuleres med korte piberensere, som hvepsene midlertidigt vil klæbe sig til.
Kvantificering af Lotka-Volterra-konkurrencemodellen:
Lotka-Volterra-konkurrencemodellen blev beskrevet og defineret med ligninger i indledningen. Som nævnt der kan ligningen, hvis værdierne for hver ligning kan estimeres empirisk ud fra resultaterne af et forsøg, anvendes til at forudsige det potentielle resultat af en konkurrence (dvs. om de to arter vil sameksistere, eller om den ene i sidste ende vil udelukke den anden). Værdierne for K1, K2, alpha12 og alpha21 anvendes til at plotte isoklinerne for nulvækst (dvs. hvor dN1/dt eller dN2/dt er lig nul) for begge arter på den samme graf, og de resulterende summer af populationsvækstvektorer (baner) anvendes til at bestemme resultatet af konkurrencen.
På grundlag af vores forsøgsplan skal vi bestemme værdierne for disse parametre. Husk på, at bæreevnen for en population er det maksimale antal individer, der kan overleve i et levested. For enkelhedens skyld har vi i dette forsøg defineret parasitoidernes levested som en enkelt vært. I virkeligheden vil et levested naturligvis sandsynligvis indeholde mere end én mudderduberrede eller en puppe af en snylteflue, og der vil derfor være mange potentielt udnyttelige værter. For at bestemme de to arters bæreevne er vi nødt til at kende det maksimale antal afkom af en given art, der kan produceres på en enkelt vært, når kun den pågældende art er til stede. På denne baggrund kan data fra hvilken behandling anvendes til at vurdere bæreevnen for Melittobia og Nasonia? (Husk, at ved bæreevne vil alle værtsressourcerne blive udnyttet.)
Det er lidt mere kompliceret at estimere konkurrencekoefficienterne (alpha12 og alpha21). Husk, at ligningen for populationsvæksten for art 1 er:
Når alle værtsressourcerne er brugt af parasitoiderne, kan en population ikke længere vokse. Med andre ord, dN1 /dt = 0. Denne tilstand vil opstå, når K1 – N1 – alpha12*N2 = 0. For at finde alpha12 skal vi løse den (dvs. lave lidt algebra) og derefter erstatte værdierne for K1, N1 og N2 med værdier for K1, N1 og N2. Ovenfor beskrev vi, hvordan man finder bæreevnen. Hvis vi antager, at Melittobia er “art 1”, skal vi bruge dens bæreevne til K1. Antallet af Melittobia- og Nasonia-efterkommere, der produceres i interspecifik konkurrence, er henholdsvis N1 og N2. På denne baggrund skal data fra hvilken behandling anvendes til at anslå N1 og N2?
Den samme fremgangsmåde, som du brugte til at beregne alpha12, kan bruges til at beregne alpha21.
Nu, hvor du har beregnet alle parameterværdierne, kan du bruge disse værdier til at plotte nulvækstisoklinerne og forudsige resultatet af konkurrencen mellem Melittobia og Nasonia.
Retningslinjer for dataanalyse:
Vi kan bruge sammenligninger mellem forskellige behandlinger til at undersøge den relative betydning af intraspecifik og interspecifik konkurrence. Først skal det identificeres, hvilken type konkurrence, intraspecifik eller interspecifik, der eventuelt forekommer i hver behandling. Når du har gjort dette, skal du tænke over alle sammenligningerne mellem behandlingsparrene. Hvad fortæller hver af sammenligningerne os? Det kan være nyttigt at udarbejde et skema med en liste over sammenligningerne og deres betydning. Da alle sammenligningerne omfatter to behandlinger, kan de analyseres statistisk ved hjælp af t-tests.
Spørgsmål til videre overvejelse og diskussion:
- Baseret på de parameterværdier, som du har beregnet for Lotka-Volterra-konkurrencemodellen, hvad er det forudsagte resultat af konkurrencen mellem de to arter? Blev det forudsagte resultat opnået i hver gentagelse af den interspecifikke konkurrence? Hvis ikke, hvorfor ikke?
- “Gauses lov” siger, at konkurrenter, der deler præcis de samme ressourcer på samme måde, ikke kan eksistere side om side. Det betyder, at den art, der mest effektivt udnytter den omstridte ressource, i sidste ende vil eliminere den anden art på det pågældende sted. Ser Gause’s lov ud til at gælde for samspillet mellem Melittobia og Nasonia? Hvorfor eller hvorfor ikke?
- Hvis disse to arter skulle bruge den samme vært i naturen, hvordan kan ressourcefordelingen så gøre det muligt for dem at eksistere side om side?
- På baggrund af resultaterne af dit eksperiment, hvorfor bruger de to arter så ikke den samme vært i naturen?
- På baggrund af de anslåede værdier for bæreevne og konkurrencekoefficienter skal du forudsige resultatet af konkurrencen mellem Melittobia og Nasonia ved hjælp af Lotka-Volterra-konkurrencemodellen for Populus (se referencer og links). Påvirkes det forudsagte resultat af konkurrencen af de oprindelige populationsstørrelser eller populationsvæksthastigheder? I bekræftende fald, hvordan? Hvordan påvirkes tiden til at nå ligevægt af disse værdier?
- Bærkapaciteterne og konkurrencekoefficienterne er kun skøn. Hvilke faktorer kan påvirke bæreevnen og konkurrencekoefficienterne for disse to arter?
- Hvis der forekommer interspecifik konkurrence hos disse arter, hvordan kan vi så afgøre, hvilken konkurrencemekanisme (interferens eller udnyttelse) der forekommer?
*** Bemærk: Svar på mange af disse spørgsmål og talrige andre kommentarer fra den medvirkende forfatter kan findes i “NOTES TO FACULTY: kommentarer til spørgsmål til videre overvejelse”.
Referencer og links:
- Abe, J., Y. Kamimura, N. Kondo, og M. Shimada. 2003. Ekstremt kvindelig kønsfordeling og dødelig han-han-kamp hos en parasitoid hveps, Melittobia australica (Eulophidae). Behavioural Ecology 14: 34-39.
- Amarasekare, P. 2000. Sameksistens af konkurrerende parasitoider på en spredt spredt vært: lokale vs. rumlige mekanismer. Ecology 81: 1286-96.
- Cooperband M. F., R. W. Matthews, S. B. Vinson. 2003. Faktorer, der påvirker Melittobia digitata’s (Hymenoptera: Eulophidae) reproduktionsbiologi og manglende opfyldelse af kønsfordelingen i henhold til Hamiltons teori om lokal parringskonkurrence. Entomologia Experimentalis et Applicata 109: 1-12.
- Côsoli, F. L., og S. B. Vinson. 2002. Koblingsstørrelse, udvikling og differentiering af vingemorfologi hos Melittobia digitata. Entomologia Experimentalis et Applicata 102: 135-143.
- Freeman, B. E., og K. Ittyeipe. 1976. Feltundersøgelser af den kumulative reaktion af Melittobia sp. Hawaiiensis complex) (Eulophidae) på varierende værtstætheder. Journal of Animal Ecology 45: 415-423.
- Gause, G. F. 1934. Kampen for eksistens. Williams and Wilkins, Baltimore.
- Harden, G. 1960. Det konkurrencemæssige udelukkelsesprincip. Science 131: 1292-1297.
- Hawkins, B. A. 2000. Sameksistens af arter i parasitoid-samfund: Har konkurrence betydning? Siderne 198-213 i M. E. Hochberg og A. R. Ives (eds). Parasitoid Population Biology. Princeton University Press, Princeton, New Jersey.
- King, B. 2000. Nasonia vitripennis, juvelhvepsen. Illustrating science and biology concepts with an insect. www.bios.niu.edu/bking/nasonia.htm
- Krebs, C. J. 1994. Økologi. Den eksperimentelle analyse af udbredelse og hyppighed. 4. udgave. HarperCollins College Publishers, New York, NY.
- Mackauer, M., B. Bai, A. Chow, og T. Danyk. 1992. Asymmetrisk larvekonkurrence mellem to arter af solitære snyltehvepse: indflydelse af superparasitter. Ecological Entomology 17: 233-236.
- Price, P. 1970. Parasitoider, der udnytter den samme vært: adaptiv karakter af forskelle i størrelse og form. Ecology 53: 445-454.
- Price, P. 1972. Karakteristika, der muliggør sameksistens blandt parasitoider af en savflue i Quebec. Ecology 51: 190-195.
- Strand, M. R. 2002. Interaktioner mellem parasitoider i larvestadiet og deres værter. Pages 129-152 in E. E. Lewis, J. F. Campbell, and M. V. K. Sukhdeo (eds). The Behavioural Ecology of Parasites. CABI Publishing, New York.
- Silva-Torres, C., og R. W. Matthews. 2003. Udvikling af Melittobia australica Girault og M. digitata Dahms (Hymenoptera: Eulophidae), der parasiterer Neobellieria bullata (Parker) (Diptera: Sarcophagidae) pupper. Neotropical Entomology 32: 645-651.
- Werren, J. H. 1983. Kønskvoteudvikling under lokal parringskonkurrence hos en parasitær hveps. Evolution 37:116-124.
- Werren, J.H. 1984. Regulering af ynglestørrelse og kønskvotient hos den parasitiske hveps Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae). Netherlands Journal of Zoology 34: 123-143.
For oplysninger om naturhistorie og levesteder, se:
- www.wowbugs.com for Melittobia,
www.bios.niu.edu/bking/nasonia.htm for Nasonia,
www.rochester.edu/College/BIO/labs/WerrenLab/nasonia/ for Nasonia.
Populus kan downloades fra www.cbs.umn.edu/populus.
Tools for Assessment of Student Learning Outcomes:
Evalueringen er blevet foretaget på forskellige måder på de forskellige institutioner, der har anvendt denne øvelse. I alle tilfælde evalueres de studerende på grundlag af en videnskabelig artikel, som hver studerende har skrevet individuelt eller i en gruppe. I nogle tilfælde er de studerende blevet evalueret på både første og andet udkast af en opgave.
Rubrikken for bedømmelsen af opgaverne varierer fra underviser til underviser. Nedenfor er et eksempel på en pointrubrik, der anvendes på Morehouse College for en “resultatsammenfatning”, som indeholder alle komponenterne i en videnskabelig artikel undtagen metoderne. I denne evalueringsrubrik vedrører “målgruppe” den studerendes valg af passende målgruppe. Det forventes, at eleverne skriver deres rapport, som om det var en videnskabelig artikel. Den passende målgruppe er således en gruppe af jævnaldrende, som ikke har udført forsøget, men som har videnskabelige færdigheder. Rapporter, der er skrevet til underviseren eller til andre studerende i klassen, har ikke den rette målgruppe. “Format” er rapportens overordnede organisering i afsnit, der har en parallel organisering og bygger på hinanden. For eksempel skal diskussionen evaluere de resultater, der er rapporteret i resultaterne, og sætte disse resultater ind i en større sammenhæng. Diskussionen bør også omhandle den hypotese, der er anført i indledningen.
| Sammenfattende evaluering af resultaterne (50 mulige point) | |
| Indledning og titelside (10 point) | _____ |
| Resultater (10 point) | _____ |
| Diskussion og konklusioner (10 point) | _____ |
| Litteraturanvendelse og citater (10 point) | _____ |
| Format, Målgruppe (10 point) | _____ |
På det indledende niveau afleverer eleverne alle klassens tabeller, svarene på diskussionsspørgsmålene og ugentlige notater om undersøgelsens forløb. Ud over individuelle videnskabelige artikler kan eleverne præsentere resultaterne af forsøget i form af videnskabelige artikler i grupper, mundtlige præsentationer i grupper eller plakatpræsentationer i grupper. Men da alle eleverne udfører det samme eksperiment og derfor præsenterer de samme resultater, vil individuelle videnskabelige artikler eller gruppearbejder være de mest effektive.
Værktøjer til formativ evaluering af dette forsøg:
I økologikurset på Morehouse College bliver hver studerende bedt om at evaluere hver laboratorieøvelse på en 10-punktsskala med hensyn til den opfattede effektivitet af hver undersøgelse i forhold til at styrke deres viden om og forståelse af de emner, der er behandlet i forelæsningskurset i økologi. Desuden spørges de studerende om, hvilke øvelser der var mindst og mest fornøjelige, og hvilke øvelser der øgede deres forståelse af den videnskabelige metode mest. Disse oplysninger bruges til at vælge og ændre de øvelser, der anvendes i de efterfølgende semestre.
I økologikurserne på Emory University og Radford University bliver hver enkelt studerende spurgt, hvilke øvelser de kunne lide bedst og hvilke de kunne lide mindst. De studerende giver undertiden kommentarer til, hvorfor de vurderede øvelserne på den måde, som de gjorde. Disse oplysninger bruges igen til at ændre de øvelser, der anvendes i de efterfølgende semestre.
En omfattende diskussion om evaluering findes i afsnittet om undervisning på dette websted.