
Escribe aquí tus ejemplos.
- 9.1 Moléculas de señalización y receptores celulares
- 9.2 Propagación de la señal
- 9.3 Respuesta a la señal
Imagina cómo sería la vida si tú y las personas que te rodean no pudieran comunicarse. No podrías expresar tus deseos a los demás, ni podrías hacer preguntas para saber más sobre tu entorno. La organización social depende de la comunicación entre los individuos que componen esa sociedad.
Al igual que ocurre con las personas, es vital que las células individuales puedan interactuar con su entorno y entre sí. Esto es así tanto si una célula crece sola en un estanque como si es una de las muchas células que forman un organismo mayor. Para responder adecuadamente a los estímulos externos, las células han desarrollado complejos mecanismos de comunicación para poder recibir un mensaje, transferir la información a través de la membrana plasmática y, a continuación, producir cambios dentro de la célula en respuesta al mensaje.
En los organismos multicelulares, las células envían y reciben constantemente mensajes químicos para coordinar las acciones de otros órganos, tejidos y células. La capacidad de enviar mensajes de forma rápida y eficaz permite a las células coordinar y ajustar sus funciones. Aunque la necesidad de comunicación celular en los organismos más grandes parece obvia, incluso los organismos unicelulares se comunican entre sí. Las células de las levaduras se comunican entre sí para facilitar el apareamiento. Algunas formas de bacterias coordinan sus acciones para formar grandes complejos llamados biofilms o para organizar la producción de toxinas para eliminar organismos competidores. La capacidad de las células para comunicarse mediante señales químicas se originó en las células individuales y fue esencial para la evolución de los organismos multicelulares. La comunicación eficiente y sin errores es vital para toda la vida.
- 9.1 | Moléculas de señalización y receptores celulares
- 9.1.1 Formas de señalización
- Señalización paracrina
- Señalización autocrina
- Señalización endocrina
- Señalización directa
- 9.1.2 Tipos de receptores
- Receptores internos
- Receptores de la superficie celular
- 9.1.3 Moléculas señalizadoras
- Ligandos hidrofóbicos pequeños
- Ligandos solubles en agua
- 9.2 | Propagación de la señal
- 9.2.1 Vías de señalización y amplificación de señales
- 9.2.2 Métodos de señalización intracelular
- Fosforilación
- Segundos mensajeros
- 9.3 | Respuesta a la señal
- 9.3.1 Respuestas a la vía de señalización
- Expresión génica
- Aumento del metabolismo celular
- Crecimiento celular
- Muerte celular
- 9.3.2 Terminación de las vías de señalización
9.1 | Moléculas de señalización y receptores celulares
Al finalizar esta sección, será capaz de:
- Describir cuatro tipos de señalización que se encuentran en los organismos multicelulares.
- Comparar los receptores internos con los receptores de la superficie celular.
- Reconocer la relación entre la química de un ligando y su mecanismo de acción.
Hay dos tipos de comunicación en el mundo de las células vivas. La comunicación entre células se llama señalización intercelular, y la comunicación dentro de una célula se llama señalización intracelular. Una forma fácil de recordar la distinción es entender que el prefijo inter- significa «entre» (una autopista interestatal cruza entre estados) e intra- significa «dentro» (un IV significa intravenoso o «dentro de la vena»).
Las señales químicas son liberadas por una célula señalizadora y recibidas por una célula objetivo. Las células diana tienen proteínas denominadas receptores, que se unen a las moléculas señalizadoras y provocan una respuesta. Las moléculas señalizadoras que se unen a los receptores se denominan ligandos. Los ligandos y los receptores son específicos entre sí; un receptor suele unirse sólo a su ligando específico. Sin embargo, hay diferentes tipos de señalización.
9.1.1 Formas de señalización
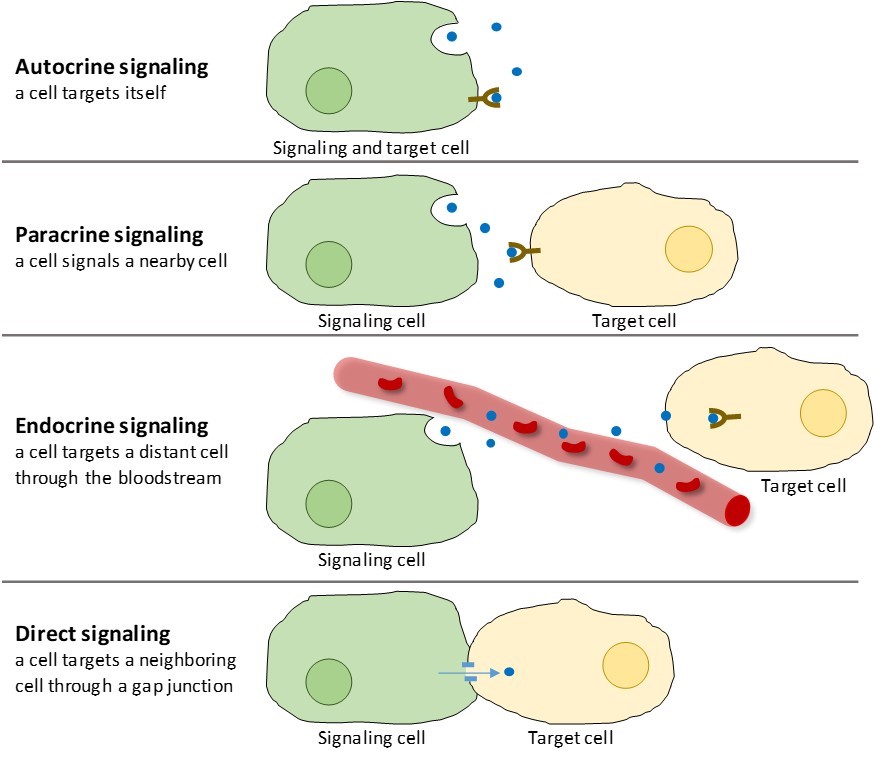
Hay cuatro categorías de señalización química que se encuentran en los organismos multicelulares: señalización autocrina, señalización paracrina, señalización endocrina y señalización directa a través de las uniones en hueco (figura 9.2). La principal diferencia entre las distintas categorías de señalización es la distancia que recorre la señal para llegar a la célula diana.
Señalización paracrina
Las señales que actúan localmente entre células que están próximas se denominan señales paracrinas. Las señales paracrinas se mueven por difusión a través de la matriz extracelular (Figura 9.2). Este tipo de señales suele provocar respuestas rápidas que duran poco tiempo. Para mantener la respuesta localizada, los ligandos paracrinos suelen ser rápidamente degradados por enzimas o eliminados por las células vecinas. La eliminación de las señales restablece el gradiente de concentración para la molécula de señal, lo que permite que se difundan rápidamente por el espacio intracelular si se liberan de nuevo.
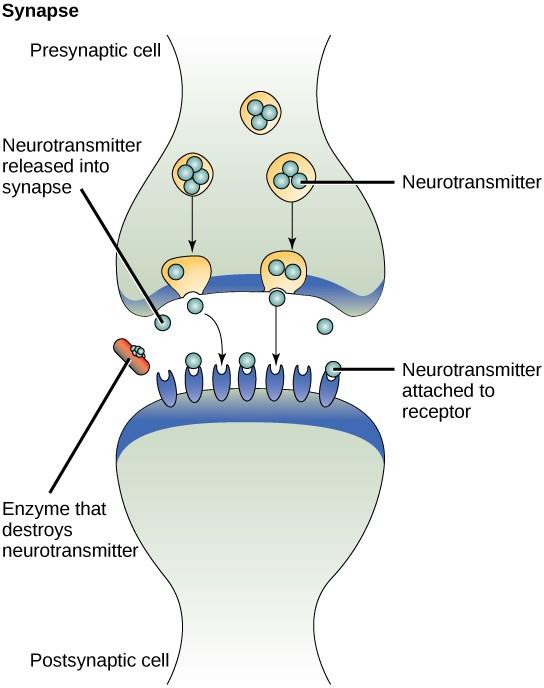
Un ejemplo de señalización paracrina es la transferencia de señales entre células nerviosas. El pequeño espacio entre las células nerviosas donde se produce la transmisión de señales se denomina sinapsis. Las señales se propagan a lo largo de las células nerviosas mediante impulsos eléctricos rápidos. Cuando estos impulsos llegan al final de una célula nerviosa, la célula presináptica (la que emite la señal) libera en la sinapsis unos ligandos químicos llamados neurotransmisores. Los neurotransmisores se difunden a través de la sinapsis (Figura 9.3). La pequeña distancia entre las células nerviosas permite que la señal se desplace rápidamente, lo que permite una respuesta inmediata, como por ejemplo, «¡quita la mano de la estufa!». Cuando el neurotransmisor se une al receptor en la superficie de la célula postsináptica, se lanza el siguiente impulso eléctrico. Los neurotransmisores se degradan rápidamente o son reabsorbidos por la célula presináptica para que la célula nerviosa receptora pueda recuperarse rápidamente y estar preparada para responder rápidamente a la siguiente señal sináptica.
Señalización autocrina
Cuando una célula responde a su propia molécula de señalización, se denomina señalización autocrina (auto = «propio»). La señalización autocrina se produce a menudo con otros tipos de señalización. Por ejemplo, cuando se libera una señal paracrina, la célula señalizadora puede responder a la señal junto con sus vecinas (Figura 9.2).
La señalización autocrina se produce a menudo durante el desarrollo temprano de un organismo para asegurar que las células se desarrollen en los tejidos correctos. La señalización autocrina también regula la sensación de dolor y las respuestas inflamatorias. Además, si una célula está infectada con un virus, la célula puede señalarse a sí misma para someterse a la muerte celular programada, matando al virus en el proceso.
Señalización endocrina
Las señales de las células distantes se llaman señales endocrinas, y se originan en las células endocrinas. (En el cuerpo, muchas células endocrinas se encuentran en las glándulas endocrinas, como la glándula tiroides, el hipotálamo y la hipófisis). Este tipo de señales suelen producir una respuesta más lenta pero tienen un efecto más duradero. Los ligandos liberados en la señalización endocrina se denominan hormonas, moléculas señalizadoras que se producen en una parte del cuerpo pero que afectan a otras regiones corporales a cierta distancia (figura 9.2).
Las hormonas recorren las grandes distancias entre las células endocrinas y sus células diana a través del torrente sanguíneo, que es una forma relativamente lenta de desplazarse por el cuerpo. Debido a su forma de transporte, las hormonas se diluyen y están presentes en bajas concentraciones cuando actúan sobre sus células diana. Esto es diferente de la señalización paracrina, en la que las concentraciones locales de las moléculas señalizadoras pueden ser muy elevadas.
Señalización directa
Las uniones en hueco en los animales y los plasmodesmos en las plantas son conexiones entre las membranas plasmáticas de las células vecinas. Estos canales llenos de agua permiten la difusión de pequeñas moléculas de señalización entre las dos células. Las moléculas pequeñas, como los iones de calcio (Ca2+), pueden moverse entre las células, pero las moléculas grandes, como las proteínas y el ADN, no pueden pasar por los canales. La especificidad de los canales garantiza que las células sigan siendo independientes, pero que puedan transmitir señales con rapidez y facilidad. La señalización directa permite a un grupo de células coordinar su respuesta a una señal que sólo una de ellas puede haber recibido. En las plantas, los plasmodesmos son ubicuos, lo que convierte a toda la planta en una gigantesca red de comunicación.
9.1.2 Tipos de receptores
Los receptores son moléculas proteicas en la célula diana o en su superficie que se unen a ligandos. Existen dos tipos de receptores, los receptores internos y los receptores de la superficie celular.
Receptores internos
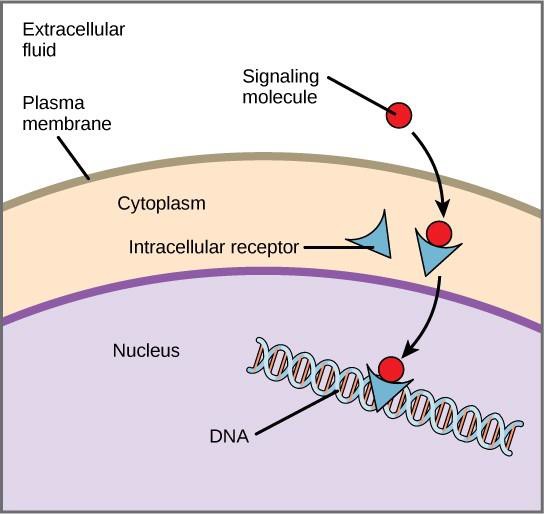
Los receptores internos, también conocidos como receptores intracelulares o citoplasmáticos, se encuentran en el citoplasma de las células diana y responden a moléculas de ligandos hidrofóbicos que son capaces de atravesar la membrana plasmática. Una vez en el interior de la célula, muchas de estas moléculas se unen a proteínas que actúan como reguladores de la síntesis del ARNm (transcripción) para mediar en la expresión de los genes.
La expresión de los genes es el proceso celular de transformación de la información del ADN de una célula en una secuencia de aminoácidos, que finalmente forma una proteína. Cuando el ligando se une al receptor interno, se desencadena un cambio conformacional que expone un sitio de unión al ADN en la proteína del receptor. El complejo ligando-receptor se desplaza al núcleo, se une a regiones reguladoras específicas del ADN cromosómico y promueve el inicio de la transcripción (figura 9.4). La transcripción es el proceso de copiar la información del ADN de una célula en una forma especial de ARN llamada ARN mensajero (ARNm); la célula utiliza la información del ARNm para unir aminoácidos específicos en el orden correcto, produciendo una proteína. Así, cuando un ligando se une a un receptor interno, puede influir directamente en la expresión génica de la célula diana.
Receptores de la superficie celular
Los receptores de la superficie celular, también conocidos como receptores transmembrana, son proteínas integrales que se unen a moléculas de señalización externas. Estos receptores abarcan la membrana plasmática y realizan la transducción de señales, en la que una señal extracelular se convierte en una señal intercelular. (Figura 9.5). Dado que las proteínas receptoras de la superficie celular son fundamentales para el funcionamiento normal de la célula, no debe sorprender que un mal funcionamiento de cualquiera de estas proteínas pueda tener graves consecuencias. Se ha demostrado que los errores en las estructuras proteicas de ciertas moléculas receptoras desempeñan un papel en la hipertensión (presión arterial alta), el asma, las enfermedades cardíacas y el cáncer.
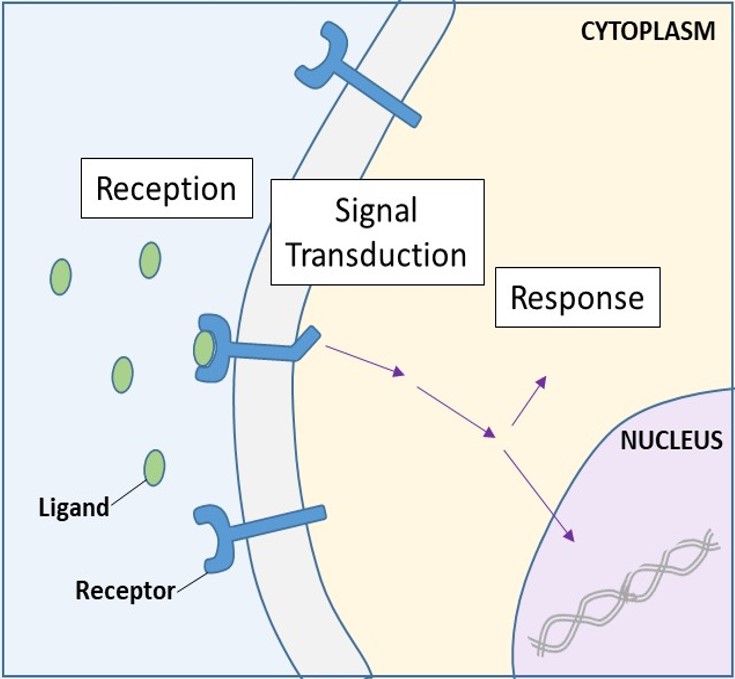
Cada receptor de la superficie celular tiene tres componentes principales: un dominio externo de unión al ligando, o dominio extracelular; una región hidrofóbica que atraviesa la membrana; y un dominio intracelular. Los receptores de la superficie celular están implicados en la mayor parte de la señalización en los organismos multicelulares. Existen tres categorías generales de receptores de superficie celular: receptores ligados a enzimas, receptores ligados a canales iónicos y receptores ligados a proteínas G.
Los receptores ligados a enzimas son receptores de superficie celular con dominios intracelulares que están asociados a una enzima. En algunos casos, el propio dominio intracelular del receptor es una enzima. Otros receptores ligados a enzimas tienen un pequeño dominio intracelular que interactúa directamente con una enzima. Los receptores ligados a enzimas normalmente tienen grandes dominios extracelulares e intracelulares, pero la región que abarca la membrana consiste en una única hélice alfa en la cadena peptídica.
Cuando un ligando se une al dominio extracelular de un receptor ligado a enzimas, se transfiere una señal a través de la membrana, activando la enzima. La activación de la enzima desencadena una cadena de acontecimientos dentro de la célula que finalmente conduce a una respuesta.
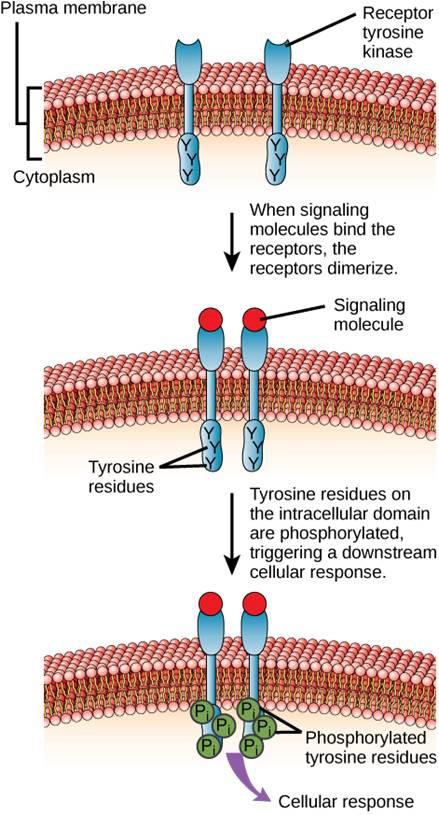
Un ejemplo de receptor ligado a una enzima es el receptor tirosina quinasa (Figura 9.6). Una cinasa es una enzima que transfiere grupos fosfato del ATP a otra proteína. El receptor de la tirosina quinasa transfiere grupos fosfato a las moléculas de tirosina. Primero, las moléculas de señalización se unen al dominio extracelular de dos receptores tirosina quinasa cercanos. A continuación, los dos receptores vecinos se unen, o dimerizan. A continuación, se añaden fosfatos a los residuos de tirosina del dominio intracelular de los receptores (fosforilación). Los residuos fosforilados pueden entonces transmitir la señal al siguiente mensajero dentro del citoplasma.
Los receptores del factor de crecimiento epidérmico son un ejemplo de receptores tirosina quinasa que siguen este modo de señalización. Los defectos en la señalización ErbB de esta familia pueden dar lugar a enfermedades neuromusculares como la esclerosis múltiple y la enfermedad de Alzheimer.
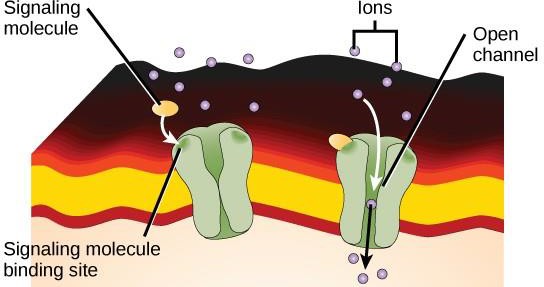
Los receptores vinculados a canales iónicos se unen a un ligando y abren un canal a través de la membrana que permite el paso de iones específicos. Este tipo de receptor de la superficie celular tiene una extensa región que abarca la membrana con aminoácidos hidrofóbicos. En cambio, los aminoácidos que recubren el interior del canal son hidrofílicos para permitir el paso de los iones. Cuando un ligando se une a la región extracelular del canal, se produce un cambio conformacional en la estructura de la proteína que permite el paso de iones como el sodio, el calcio, el magnesio o el hidrógeno (figura 9.7).
Los receptores ligados a la proteína G se unen a un ligando y activan una proteína G asociada. La proteína G activada interactúa entonces con una proteína de membrana cercana, que puede ser un canal iónico o una enzima (Figura 9.8). Todos los receptores ligados a la proteína G tienen siete dominios transmembrana, pero cada receptor tiene un dominio extracelular específico y un sitio de unión a la proteína G.
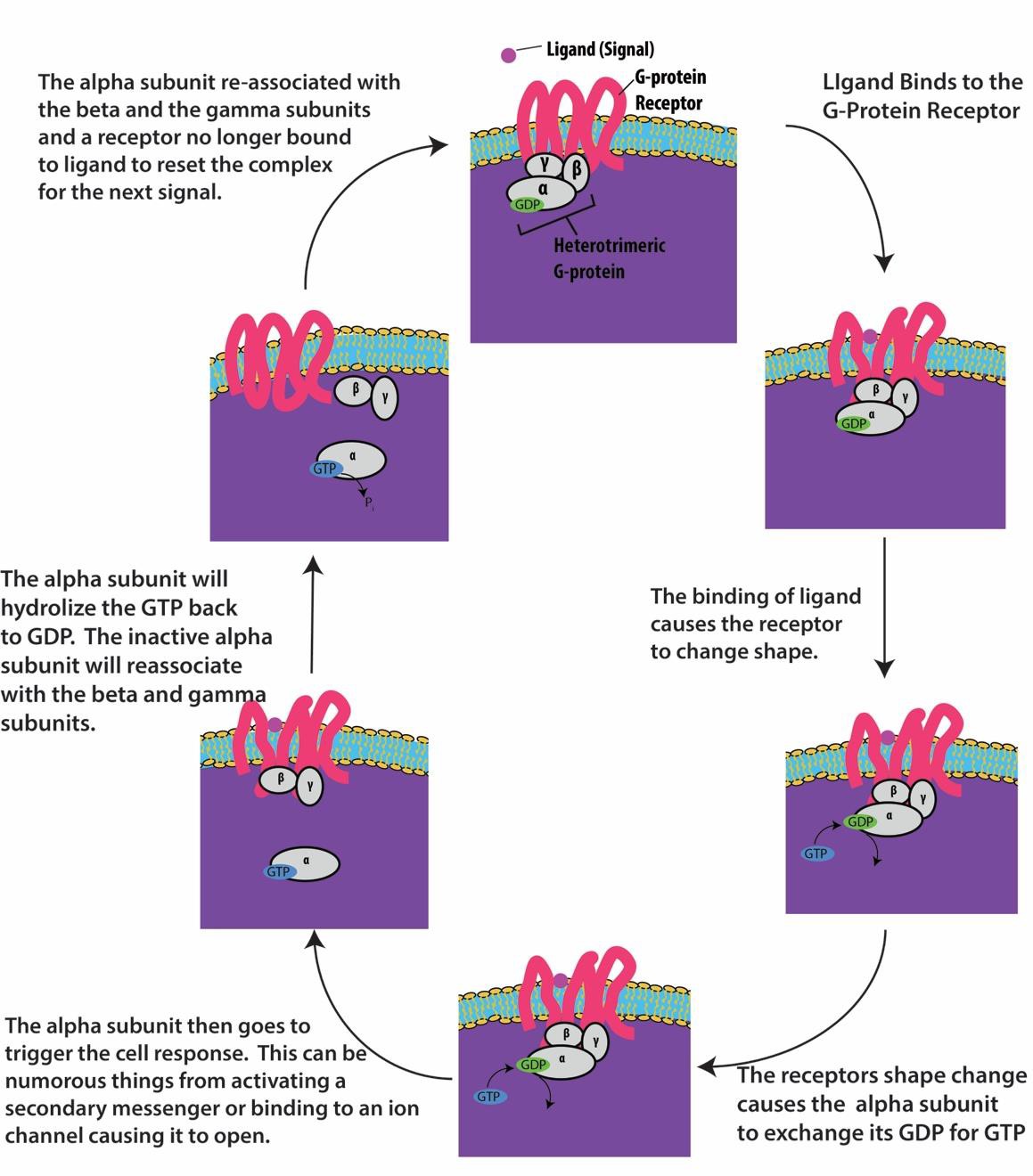
La señalización celular mediante receptores ligados a la proteína G se produce como un ciclo. Una vez que el ligando se une al receptor, el cambio de forma resultante activa la proteína G, que libera el GDP y recoge el GTP. Las subunidades de la proteína G se dividen entonces en subunidades α y βγ. Uno o ambos fragmentos de la proteína G pueden activar otras proteínas de la célula. Al cabo de un tiempo, el GTP de la subunidad α activa de la proteína G se hidroliza a GDP y la subunidad βγ se desactiva. Las subunidades se vuelven a asociar para formar la proteína G inactiva y el ciclo comienza de nuevo (figura 9.8).
Los receptores unidos a la proteína G se utilizan en muchos procesos fisiológicos, incluidos los de transducción de la visión, el gusto y la regulación del sistema inmunitario y la inflamación.

HER2 es un receptor tirosina quinasa. En el 30 por ciento de los cánceres de mama humanos, el HER2 está permanentemente activado, lo que da lugar a una división celular no regulada. El lapatinib, un fármaco utilizado para tratar el cáncer de mama, inhibe el proceso por el que el receptor se fosforila a sí mismo, reduciendo así el crecimiento del tumor en un 50 por ciento. ¿Cuál de los siguientes pasos sería inhibido por Lapatinib?
- Unión de la molécula de señalización, dimerización, y la respuesta celular posterior
- Dimerización, y la respuesta celular posterior
- La respuesta celular posterior
- Actividad de la fosfatasa, dimerización, y la respuesta celular posterior
9.1.3 Moléculas señalizadoras
Producidas por las células señalizadoras, los ligandos son señales químicas que viajan hasta las células diana y provocan una respuesta. Los tipos de moléculas que sirven como ligandos son increíblemente variados y van desde pequeñas proteínas hasta pequeños iones. Los ligandos se clasifican en ligandos hidrofóbicos pequeños, que pueden atravesar las membranas plasmáticas, o ligandos hidrosolubles, que no pueden hacerlo.
Ligandos hidrofóbicos pequeños
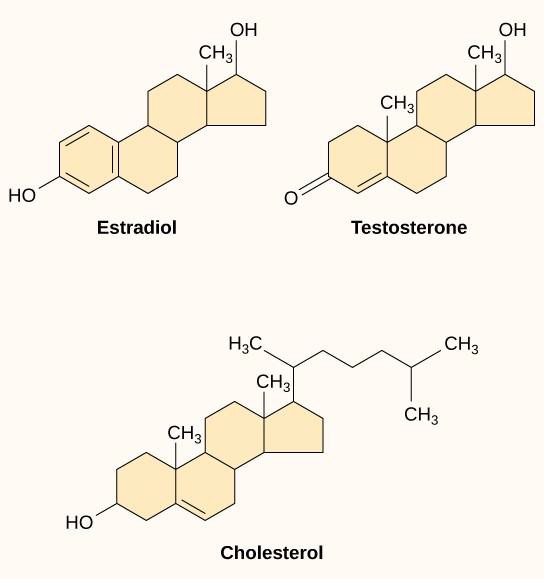
Los ligandos hidrofóbicos pequeños, también llamados ligandos liposolubles, pueden difundirse directamente a través de la membrana plasmática e interactuar con los receptores internos. Miembros importantes de esta clase de ligandos son las hormonas esteroides. Los esteroides son lípidos que tienen un esqueleto de hidrocarburos con cuatro anillos fusionados; diferentes esteroides tienen diferentes grupos funcionales unidos al esqueleto de carbono. Las hormonas esteroides incluyen la hormona sexual femenina estradiol, que es un tipo de estrógeno; la hormona sexual masculina testosterona; y el colesterol, que es un importante componente estructural de las membranas biológicas y un precursor de las hormonas esteroides (Figura 9.10). Otras hormonas hidrofóbicas son las hormonas tiroideas y la vitamina D. Para ser solubles en la sangre, los ligandos hidrofóbicos deben unirse a las proteínas transportadoras mientras se transportan por el torrente sanguíneo.
Ligandos solubles en agua
Como los ligandos solubles en agua son polares, no pueden atravesar la membrana plasmática sin ayuda. A veces son demasiado grandes para atravesar la membrana. En cambio, la mayoría de los ligandos hidrosolubles se unen al dominio extracelular de los receptores de la superficie celular (véase la figura 9.5). Este grupo de ligandos es bastante diverso e incluye moléculas pequeñas, péptidos y proteínas.
9.2 | Propagación de la señal
Al final de esta sección, será capaz de:
- Explicar cómo la unión de un ligando inicia la transducción de la señal en toda la célula.
- Reconocer que las señales intracelulares se transmiten mediante el papel de la fosforilación o de los segundos mensajeros.
Una vez que un ligando hidrosoluble se une a su receptor, la señal se transmite a través de la membrana y hacia el citoplasma. La continuación de una señal de esta manera se denomina transducción de señales (Figura 9.5). La transducción de la señal sólo se produce con los receptores de la superficie celular, ya que los receptores internos pueden entrar en la célula.
Cuando un ligando se une a su receptor, se producen cambios conformacionales que afectan al dominio intracelular del receptor. Estos cambios conformacionales conducen a la activación del dominio intracelular o de sus proteínas asociadas. En algunos casos, la unión del ligando provoca la dimerización del receptor, lo que significa que dos receptores se unen entre sí para formar un complejo estable llamado dímero. Un dímero es un compuesto químico que se forma cuando dos moléculas (a menudo idénticas) se unen. La unión de los receptores de esta manera permite que sus dominios intracelulares entren en estrecho contacto y se activen mutuamente.
9.2.1 Vías de señalización y amplificación de señales
Aunque las moléculas señalizadoras suelen encontrarse a concentraciones muy bajas, pueden producir efectos profundos. Una vez que el ligando se une al receptor de la superficie celular, la activación de los componentes intracelulares del receptor pone en marcha una cadena de acontecimientos que se denomina vía de señalización o cascada de señalización. En una vía de señalización, los segundos mensajeros, las enzimas y/o las proteínas activadas activan otras proteínas o mensajeros (figura 9.11). Cada miembro de la vía puede activar miles de veces al siguiente miembro de la vía en un proceso llamado amplificación de la señal. Dado que la señal se amplifica en cada paso, puede generarse una respuesta muy grande a partir de un solo receptor que se una a un ligando.
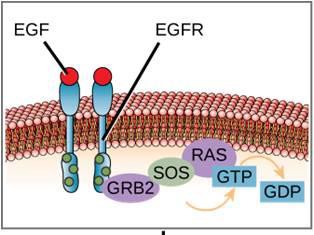
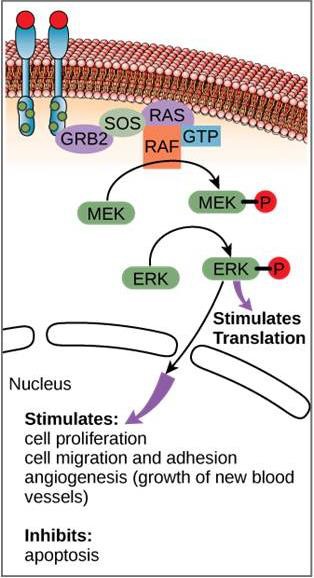
En la figura 9.11 se muestra un ejemplo de vía de señalización. El factor de crecimiento epidérmico (EGF) es una molécula de señalización que participa en la regulación del crecimiento celular, la curación de heridas y la reparación de tejidos. El receptor del EGF (EGFR) es una tirosina quinasa. Una quinasa activada fosforila y activa muchas moléculas posteriores. Cuando el EGF se une al EGFR, una cascada de eventos de fosforilación posteriores indica a la célula que crezca y se divida. Si el EGFR se activa en momentos inadecuados, puede producirse un crecimiento celular incontrolado (cáncer).
En ciertos cánceres, la actividad GTPasa de la proteína G RAS está inhibida. Esto significa que la proteína RAS ya no puede hidrolizar GTP en GDP. ¿Qué efecto tendría esto en los eventos celulares posteriores?
9.2.2 Métodos de señalización intracelular
La inducción de una vía de señalización depende de la modificación de un componente celular por una enzima. Hay numerosas modificaciones enzimáticas que pueden ocurrir para activar el siguiente componente de la vía. Los siguientes son algunos de los eventos más comunes en la señalización intracelular.
Fosforilación
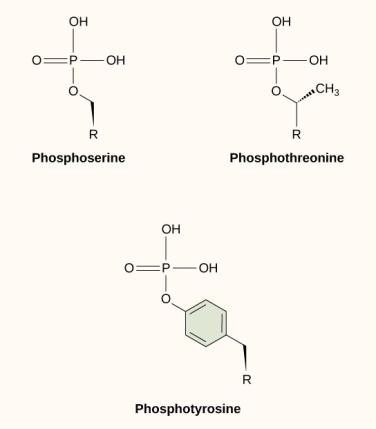
Una de las modificaciones químicas más comunes que ocurre en las vías de señalización es la adición de un grupo fosfato a una molécula en un proceso llamado fosforilación. El fosfato puede añadirse a un nucleótido como el GMP para formar GDP o GTP. Los fosfatos también se añaden a menudo a los residuos de serina, treonina y tirosina de las proteínas, donde sustituyen al grupo hidroxilo del aminoácido (Figura 9.12). La transferencia del fosfato es catalizada por una enzima llamada cinasa. La fosforilación puede activar o inactivar enzimas, y la inversión de la fosforilación, la desfosforilación, revertirá el efecto.
Segundos mensajeros
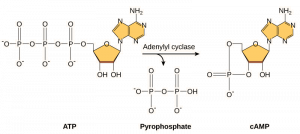
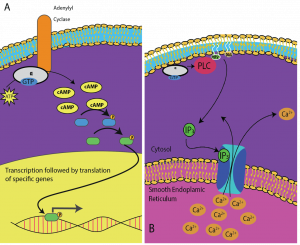
Los segundos mensajeros son pequeñas moléculas que propagan una señal después de que haya sido iniciada por la unión de la molécula de señalización al receptor. Estas moléculas ayudan a propagar una señal a través del citoplasma alterando el comportamiento de ciertas proteínas celulares. Un segundo mensajero utilizado por muchos tipos de células es el AMP cíclico (AMPc). El AMP cíclico es sintetizado por la enzima adenilil ciclasa a partir del ATP (Figura 9.13). La función principal del AMPc en las células es unirse a una enzima llamada cinasa dependiente del AMPc (A-cinasa) y activarla. La A-cinasa regula muchas vías metabólicas vitales: Fosforila residuos de serina y treonina de sus proteínas objetivo, activándolas en el proceso. La A-cinasa se encuentra en muchos tipos de células, y las proteínas objetivo en cada tipo de célula son diferentes. Otro mensajero secundario es el Ca2+que puede ser liberado para inundar la célula.
Diferentes células responden de forma diferente al AMPc. En la Figura 9.14, se muestra la subunidad alfa de un receptor de proteína G activando dos tipos diferentes de señalización. En la primera imagen, el AMPc es producido por la enzima adenilato ciclasa cuando es activada por la subunidad alfa. El AMPc activa entonces otras proteínas que afectan la transcripción de genes. En la segunda imagen, la subunidad alfa de la proteína G desencadena una cascada que libera Ca2+ del retículo endoplásmico liso. En este caso el Ca2+ es el mensajero secundario que provoca la respuesta celular.
9.3 | Respuesta a la señal
Al final de esta sección, será capaz de:
- Reconocer que las vías de señalización dirigen la expresión de proteínas, el metabolismo celular y el crecimiento celular.
- Reconocer el papel de la apoptosis en el desarrollo y mantenimiento de un organismo sano.
Utilizando las vías de transducción de señales, los receptores de la membrana plasmática producen una variedad de efectos en la célula. Dentro de la célula, los ligandos se unen a sus receptores internos, lo que les permite afectar directamente al ADN de la célula y a la maquinaria de producción de proteínas. Los resultados de las vías de señalización son muy variados y dependen del tipo de célula implicada, así como de las condiciones externas e internas. A continuación se describe una pequeña muestra de respuestas.
9.3.1 Respuestas a la vía de señalización
Expresión génica
Algunas vías de transducción de señales regulan la transcripción del ARN. Otras regulan la traducción de las proteínas.
Aumento del metabolismo celular
La activación de los receptores β-adrenérgicos en las células musculares por parte de la adrenalina conduce a un aumento del AMP cíclico en el interior de la célula. La adrenalina es una hormona producida por la glándula suprarrenal que prepara al cuerpo para emergencias a corto plazo.
Crecimiento celular
Las vías de señalización celular también desempeñan un papel importante en la división celular. Las células no se dividen normalmente a menos que sean estimuladas por señales de otras células. Los ligandos que promueven el crecimiento celular se denominan factores de crecimiento. La mayoría de los factores de crecimiento se unen a receptores de la superficie celular que están vinculados a tirosina quinasas.
Muerte celular
Cuando una célula está dañada, es superflua o potencialmente peligrosa para un organismo, una célula puede iniciar un mecanismo para desencadenar la muerte celular programada, o apoptosis. La apoptosis permite que una célula muera de una manera controlada que evita la liberación de moléculas potencialmente dañinas del interior de la célula. Sin embargo, en algunos casos, como una infección vírica o una división celular descontrolada debida al cáncer, los controles y equilibrios normales de la célula fallan. La señalización externa también puede iniciar la apoptosis. Por ejemplo, la mayoría de las células animales normales tienen receptores que interactúan con la matriz extracelular, una red de glicoproteínas que proporciona soporte estructural a las células animales. La unión de los receptores celulares a la matriz extracelular inicia una cascada de señalización dentro de la célula. Sin embargo, si la célula se aleja de la matriz extracelular, la señalización cesa y la célula se somete a la apoptosis. Este sistema ayuda a evitar que las células se desplacen por el cuerpo y proliferen sin control, como ocurre con las células tumorales que hacen metástasis.
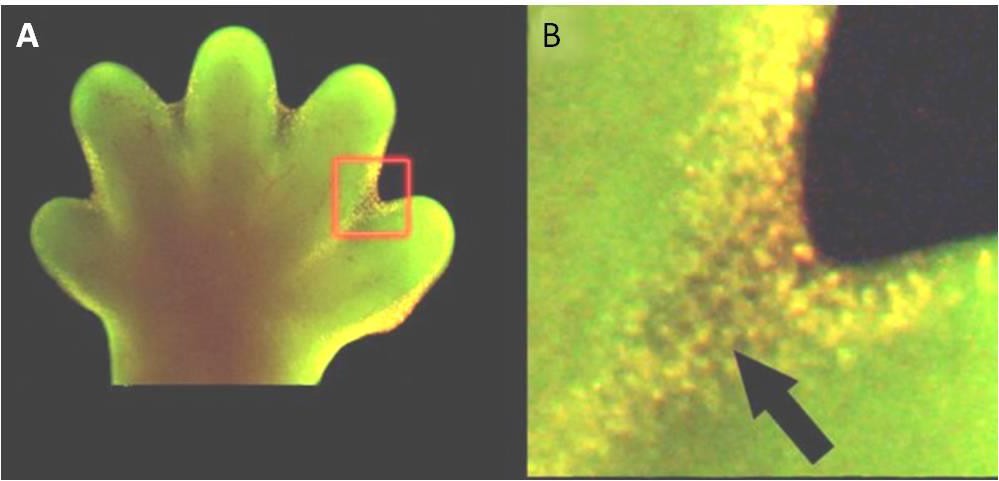
La apoptosis también es esencial para el desarrollo embriológico normal. En los vertebrados, por ejemplo, las primeras etapas del desarrollo incluyen la formación de tejido en forma de red entre los dedos individuales de las manos y los pies (Figura 9.15). En el transcurso del desarrollo normal, estas células innecesarias deben ser eliminadas, lo que permite la formación de dedos de manos y pies completamente separados. Un mecanismo de señalización celular desencadena la apoptosis, que destruye las células entre los dedos en desarrollo.
9.3.2 Terminación de las vías de señalización
La señalización aberrante que se observa a menudo en las células tumorales es una prueba de que la terminación de una señal en el momento adecuado puede ser tan importante como la iniciación de una señal. Un método para detener una señal específica es degradar el ligando o eliminarlo para que ya no pueda acceder a su receptor. Una de las razones por las que las hormonas hidrofóbicas como el estrógeno y la testosterona desencadenan eventos de larga duración es porque se unen a proteínas transportadoras. Estas proteínas permiten que las moléculas insolubles sean solubles en la sangre, pero también protegen a las hormonas de la degradación por parte de las enzimas circulantes.
Dentro de la célula, muchas enzimas diferentes revierten las modificaciones celulares que resultan de las cascadas de señalización. Por ejemplo, las fosfatasas son enzimas que eliminan el grupo fosfato unido a las proteínas por las quinasas en un proceso llamado desfosforilación. El AMPc es degradado en AMP por la fosfodiesterasa, terminando así su señal. Del mismo modo, la liberación de las reservas de calcio es revertida por las bombas de Ca2+ que se encuentran en las membranas celulares.